В современном парфюме непонятно что, а советскими можно было даже раны обработать: 10 фото легендарных одеколонов СССР
Автор Владислав На чтение 3 мин Просмотров 2.5к.
Подаренный мной отцу на День рождения парфюм стал поводом для небольшого «экскурса в историю» во время праздничного застолья.
Я только тогда узнал, что мужские парфюмы и туалетную воду советская промышленность не выпускала в принципе. Но была и альтернатива им – одеколоны. О них речь и пойдет.
Тройной одеколон
Его, если верить песне известной рок-группы, не пьет Ален Делон. Тем не менее, определенное отношение к Франции этот одеколон все же имеет.
Его появлением советские мужчины обязаны Наполеону, в свое время приказавшего создать для собственных солдат средство гигиены, одновременно обладавшее еще и лечебным, освежающим эффектами.
Стоил «Тройной» одеколон очень дешево. Спирта во флаконе содержалось примерно столько же, сколько в бутылке водки.
Использовался он и в «народной медицине» – им смазывали небольшие ссадины, ранки, прыщики, воспаления. В небольших дозах (15-20 капель на стакан воды) употребляли внутрь для лечения мигрени.
Оздоровительный эффект и характерный цитрусово-хвойный аромат обеспечивала композиция эфирных масел – шалфей, лаванда, кориандр, герань, нероли, бергамот, лимон.
Консул
Достаточно дорогой по советским меркам (4 рубля) одеколон, продукт сотрудничества советского «Союзпарфюмерпром» с французской компанией Marbel. Выпускался в Москве, на фабрике «Новая Заря».
Пик его популярности пришелся на 70-80-е годы. Аромат позиционировался как стильный и элегантный.
Начальные ноты в букете – цитрусовые (мандарин, бергамот). Далее аромат раскрывается нотами мускатного ореха, имбиря, иланг-иланга, пачули. Шлейф очень «теплый» (амбра, мускус, ладан).
В итоге получилась достаточно интересная пряно-цитрусовая композиция. Думаю, она нашла бы своих поклонников и сегодня.
Одеколон Карпаты
Выпускался на Украине, во Львове (фабрика «Красная гвоздика»). Производство началось в середине 60-х. Аромат одеколона достаточно спокойный, сдержанный, благородно-древесный.
Флакон простой, стандартной формы. Принт на этикетке и коробке одинаковый, имитирующий текстуру дерева или хвойный лес.
Создатели стремились к тому, чтобы аромат одеколона ассоциировался с Карпатами – их живописными пейзажами, величием гор, кристально чистым воздухом и реками.
Поэтому в нем отчетливо чувствуются запахи хвои и местных целебных трав. Дополнены они нежным цветочным «букетом».
Шипр
Ценился советскими мужчинами за качество и стойкость. Его производство в СССР стало возможно благодаря подарку от «братской» Индии.
Советские парфюмеры получили от коллег несколько цистерн настоящего сандалового масла, которого хватило на несколько десятилетий.
«Шипр» выпускался с середины 60-х годов на фабрике «Новая Заря». Помимо сандала, в его аромате ощущаются нотки пачули, дубового мха, бергамота.
Название и основные составляющие советские парфюмеры «позаимствовали» у француза Франсуа Коти, создавшего в 1917 году парфюм, который, по замыслу автора, должен был ассоциироваться с Кипром.
Его аромат был богатым и «аристократичным», советский вариант получился несколько проще.
О популярности «Шипра» свидетельствует то, что именно этим одеколоном клиентам освежали прическу после стрижки и умывали их после бритья. Им же были заправлены ненадолго появившиеся в Москве автоматы.
Приобрести парфюм для себя в его современном понимании могли только те советские мужчины, которые имели возможность выезжать за границу.
Абсолютному большинству граждан приходилось довольствоваться отечественными одеколонами. Использовали их в основном в санитарно-гигиенических целях, «умываясь» жидкостью после бритья.
Использовали их в основном в санитарно-гигиенических целях, «умываясь» жидкостью после бритья.
А вам приходилось пользоваться советскими мужскими одеколонами? Был особенно полюбившийся запах?
تحميل Как сделать кровь и раны в Фотошоп mp3 — mp4
Как сделать кровь и раны в Фотошоп
РЕЦЕПТ ЗОМБИ в Фотошопе Делаю Зомби из Exile в Photoshop Speed Art
Зомби апокалипсис Делаем кровь и грязь в фотошоп
How To Paint Blood In Photoshop
How To Make Fake Blood In Photoshop A Phlearn Video Tutorial
Как изменить цвет глаз в Photoshop Красные и чёрные глаза с кровью
Create Fake Blood PHOTOSHOP TUTORIAL
Гайд от Shenk II Как сделать кровь в фотошопе PS4
18 КРАФТОВЫХ ИДЕЙ С ГРИМОМ И СПЕЦЭФФЕКТАМИ
Как сделать реалестичную кровь в фотошопе
Как сделать шрам в Фотошопе
Как сделать слёзы и раны в Фотошопе
Blood Photoshop Tutorial
Tuto Photoshop Make Blood In An Eyes Color It With Chnaider Pop
Эффект шрама Как наложить фото на фото в фотошопе Photoshop 2020
Как сделать реалистичную кровь в домашних условиях DIY рецепты своими руками Дарья Дзюба
Как сделать шрам в фотошопе сS6 быстро и самое главное просто
Рисуем кровь на фотошопе Cs6
Текст из крови в фотошопе
Как превратить человека на фото в зомби
В этой статье я покажу вам, как превратить человека на фото в зомби. Вы узнаете, как использовать корректирующие слои, объединять изображения и работать с текстурами.
Вы узнаете, как использовать корректирующие слои, объединять изображения и работать с текстурами.
Программное обеспечение: Photoshop CS и выше
Ресурсы
- Модель;
- Рана;
- Мясо;
- Ржавчина 1;
- Ржавчина 2.
Сначала нам нужно подготовить фото модели. Вы должны выбрать соответствующую фотографию (например, использовать снимок человека с милой улыбкой было бы неуместно, наоборот — кричащий человек, с пронзительным взглядом подойдет намного лучше). Я использовала инструмент «Кадрирование», чтобы удалить нижнюю часть тела модели. Это позволит сфокусировать внимание на лице:
Перейдите в Слой> Новый корректирующий слой> Кривые и уменьшите яркость фото:
Используйте корректирующий слой «Цветовой тон / Насыщенность», чтобы уменьшить насыщенность цветов изображения:
Создайте новый слой (Cmd / Ctrl + Shift + N), измените для него режим смешивания на «Перекрытие», 100% и залейте слой серым цветом 50%:
Выберите инструмент «Затемнитель», установите для диапазона «Средние тона» и «Экспонирования» значения порядка 15-20%, чтобы затемнить некоторые части тела и лица модели.![]() На рисунке ниже показано, что получилось у меня при режиме смешивания «Нормальный» и при «Перекрытие»:
На рисунке ниже показано, что получилось у меня при режиме смешивания «Нормальный» и при «Перекрытие»:
Создайте новый слой, жесткой кистью с цветом #a5a39f зарисуйте глаза. На фото видно, что зрачки глаз все еще немного видны:
Я использовала корректирующий слой «Кривые» с обтравочной маской, чтобы затемнить глаза. В маске слоя я использовала мягкую кисть черного цвета, чтобы стереть области по центру глаз и придать им глубину:
Скопируйте рану из исходного изображения и поместите ее на груди модели:
Нажмите вторую справа кнопку в нижней части палитры слоев, чтобы добавить к этому слою маску. Используйте мягкую черную кисть, чтобы удалить жесткие края раны и создать плавный переход от раны к коже:
Я добавила для раны корректирующий слой «Цветовой тон / Насыщенность» и в маске этого слоя мягкой черной кистью зарисовала центр раны, чтобы сделать ее на вид не такой свежей:
Откройте текстуру «Ржавчина 1». Поместите ее под слоем модели и измените режим смешивания на «Перекрытие»:
Добавьте маску слоя и мягкой черной кистью удалите края текстуры, которые выходят за пределы тела модели:
Используйте корректирующий слой «Цветовой тон / Насыщенность», чтобы привести в соответствие эффект ржавчины с тоном кожи:
Поместите изображение «Мясо» на живот модели и измените режим смешивания этого слоя на «Умножение»:
Используйте маску слоя, чтобы смешать текстуру мяса с кожей модели:
Уменьшите интенсивность этого эффекта с помощью корректирующего слоя «Цветовой тон / Насыщенность»:
Мы продолжаем делать фигуру модели еще более «пугающей«. Откройте изображение «Ржавчина 2». Используйте инструмент «Лассо», чтобы выделить часть этой текстуры, и поместите ее на лицо модели. Установите для этого слоя режим смешивания на «Перекрытие». А также используйте маску слоя, чтобы смешать текстуру с кожей модели:
Откройте изображение «Ржавчина 2». Используйте инструмент «Лассо», чтобы выделить часть этой текстуры, и поместите ее на лицо модели. Установите для этого слоя режим смешивания на «Перекрытие». А также используйте маску слоя, чтобы смешать текстуру с кожей модели:
Возьмите другие части этой текстуры и поместите их на разные части тела модели. Вот как это сделала я:
Выберите все слои, в которых используются фрагменты текстуры «Ржавчина 2», и нажмите Cmd / Ctrl + G, чтобы сгруппировать их. Измените режим смешивания этой группы на «Перекрытие». Добавьте для нее корректирующий слой «Цветовой тон / Насыщенность», чтобы немного ослабить интенсивность эффекта ржавчины:
Используйте корректирующий слой «Кривые», чтобы затемнить этот эффект, а затем в маске этого слоя черной кистью зарисуйте те области, в которых вы не хотите затемнять ржавчину:
Мы можем добавить еще кое-что к изображению модели, чтобы сделать его еще более страшным. На новом слое мягкой кистью с цветом #940206 нарисуйте кроваво-красные участки на теле и в углу губ. Измените режим смешивания этого слоя на «Умножение»:
Измените режим смешивания этого слоя на «Умножение»:
Создайте новый слой с теми же настройками, что и в шаге 4. Используйте инструмент «Затемнитель», чтобы нарисовать морщины и вены на коже модели:
Я надеюсь, что вам понравилась эта статья:
РедакцияПеревод статьи «Turn a Portrait into a Zombie in Photoshop»
App Store: Facetune
• Facetune это забавный и мощный редактор для селфи и портретов.
———————————————————
• Лучшее приложение недели Gizmodo!
• Facetune дает каждому пользователю возможность достижения качества портретов, сравнимого только с фотошопом в руках профессионала.
Похвалы о Facetune :
• «Одно из самых мощных мобильных приложений, с которым я когда-либо сталкивался… Facetune действительно можно назвать волшебным.» — Hillel Fuld, Huffington Post
• «Я восхищена Facetune… намного ниже реальной цены.
• «легкое прикосновение, болшая разница», — Liz Gannes, AllThingsD
• «Facetune ставит возможности ретуши фотошопа на кончиках пальцев масс». — Victoria Taylor, New York Daily News
• «Трудно переоценить, насколько невероятно легко полностью изменить фотографию … к чему-то, что выглядит как принадлежащее журналу мод.» — Chris Armstrong, The Industry
———————————————————
Каждую фотографию можно подправить. Для того чтобы каждый выглядел лучшем образом, журналы используют специалистов фотошопа. До сегодняшнего дня, у всех у нас не было такой возможности. Это закончилось. Facetune обеспечивает легкие и мощные инструменты которые ранее были только у профессионалов. Теперь вы можете быть уверенны, что все ваши фотографии будут выглядет так, как вы желаете. В мире который постоянно становится все более визуальным, получить совершенную фотографию становится также важно, как причесаться перед выходом из дома!
———————————————————
С помощью Facetune вы можете:
ИДЕАЛЬНАЯ УЛЫБКА
• Подчеркнуть или расширить улыбку
• Придать зубам яркий и естественный блеск
КРАСИВАЯ КОЖА
• Разгладить морщинки и омолодить кожу
• Удалить временные недостатки, такие как прыщи и пятна
• Осветлить темные круги под глазами
ПРОНИЦАТЕЛЬНЫЕ ГЛАЗА
• Подчеркнуть глаза и создать проницательный взгляд
• Изменить цвет глаз
• Убрать эффект красного или белого глаза
ПАРИКМАХЕРСКАЯ
• Повернуть время вспять и убрать седые волосы
• Заполнить залысины
• Удалить ненужные волосы
ИЗМЕНИТЬ СТРУКТУРУ ЛИЦА
• Уточнить линии челюсти
• Приподнять скулы и брови
• Измененить форму носа
• Полностью изменить форму лица для забавного эффекта
МАКИЯЖ
• Нанести любой оттенок румян и теней для век
• Добавить объем ресницам и форму бровям
• Придать интенсивность цвету губ
ФОТО ЭФФЕКТЫ
• Расфокусировать фон, чтобы фокус был на вас
• Улучшить освещение или добавить специальные эффекты
• Добавить текстуры и рамки
• Использовать мощные локальные фильтры
ПОКАЖИТЕСЬ!
• Мгновенно послать отредактированную фотографию друзьям и семье через социальные сети или по электронной почте
ЛЕГКО И ВЕСЕЛО
• Сравнить вашу работу с оригиналом на каждом этапе
• Возникают проблемы? Facetune предлагает обширную графическую и видео помощь для каждой функции
РЕЗОЛЮЦИЯ
• iPhone 5, 5C, 5S: 12. 6 MP
6 MP
• iPhone 4S: 8 MP
• iPhone 4: 4.1 MP
Не ждите! Скачайте Facetune и создавайте впечатляющие портретные фотографии уже сегодня!
Вакансия ИБП РАН — www.ibp-ran.ru
8 февраля, 2016
Дирекция ИБП РАН объявляет конкурс на замещение вакантной должности СТАРШЕГО НАУЧНОГО СОТРУДНИКА (КАНДИДАТА НАУК) по специализации в области биоинформатики.
ДОЛЖНОСТНЫЕ ОБЯЗАННОСТИ
- Проводит научные исследования и разработки по отдельным разделам (этапам) проектов, тем в качестве ответственного исполнителя, и (или) самостоятельно осуществляет сложные исследования, эксперименты и наблюдения.
- Собирает, обрабатывает, анализирует и обобщает результаты экспериментов и наблюдений с учетом отечественных и зарубежных данных по теме исследования.
- Участвует в разработке планов и методических программ исследований, рекомендаций по использованию их результатов, а также в их практической реализации.

- Участвует в образовательном процессе (проведение семинаров и практикумов, руководство дипломными и курсовыми работами).

Должен знать: цели и задачи проводимых исследований и разработок, отечественную и зарубежную информацию по теме исследования; современные методы организации исследований, обобщения и обработки полученной информации; наблюдений; средства проведения экспериментов и наблюдений; внутренние нормативные акты, приказы и распоряжения; правила и нормы охраны: труда, пожарной безопасности.
ПРОФЕССИОНАЛЬНЫЕ НАВЫКИ: знание математических программ Origin‚ MathCAD, программ молекулярной визуализации (PyMol)‚ различных пакетов биоинформатических программ, программ по работе с векторной графикой и изображениями (Chemical Drawing programs‚ Adobe Illustrator‚ Adobe Photoshop), языка программирования R для статистической обработки данных и работы с графикой, высокоуровневого языка программирования Python.
ЛАБОРАТОРНЫЕ НАВЫКИ: электрофорез‚ хроматография, подготовка образцов для АСМ, выделение плазмидной ДНК.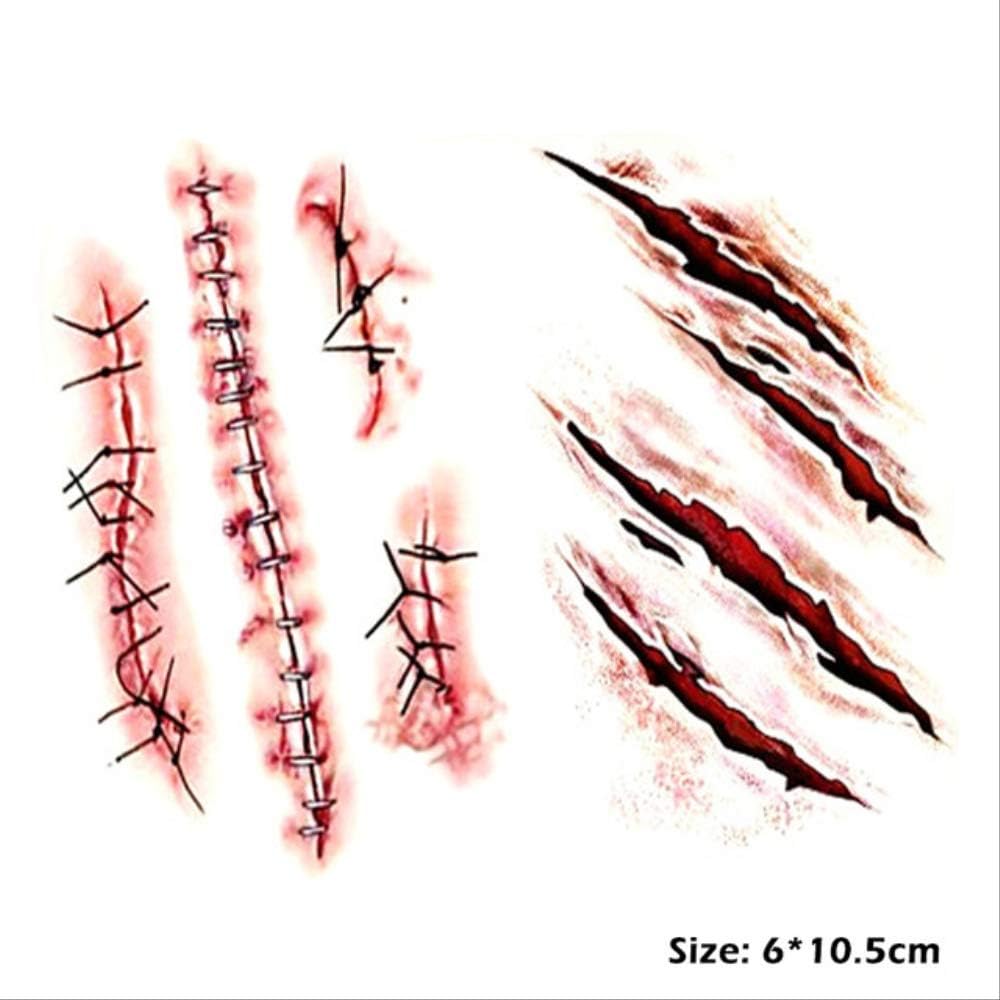
ТРЕБОВАНИЯ К КВАЛИФИКАЦИИ
- Выпускник физического факультета университета со специализацией в области биоинформатики.
- Знание принципов структурной организации белков и нуклеиновых кислот, основных биоинформатических методов их исследования.
- Опыт работы с базами данных нуклеотидных и аминокислотных последовательностей (PDB, GenBank, BLAST и др.) и статистическими методами обработки данных.
- Знание хотя бы одного современного языка программирования.
ДЛЯ УЧАСТИЯ В КОНКУРСЕ В ДИРЕКЦИЮ ИБП РАН ПОДАЮТСЯ
- Заявление в конкурсную комиссию с просьбой допустить к участию в конкурсе на должность старшего научного сотрудника.
- Отзыв (характеристика) с последнего места работы, подписанный уполномоченным работодателем должностным лицом или руководителем подразделения.
- Список трудов (статей, глав в монографиях и т.п.).
- Сведения о личном участии в научных мероприятиях (съезды, конференции, симпозиумы и иные научные мероприятия) с указанием статуса доклада (устный или стендовый).
- Данные о цитировании работ претендентов (можно привести регистрационные данные в Web of Science, ResearcherID, РИНЦ и т.п.).

Срок подачи документов — два месяца со дня публикации. Документы направляются по адресу: 142290, Московская обл., г. Пущино, ул. Институтская, 7, Федеральное государственное учреждение науки Института биологического приборостроения с опытным производством РАН. Телефон для справок: 8 (4967) 31-89-32.
назадСписок отечественных книг, поступивших с 20 февраля по 4 марта 2012 г.
…
Список отечественных книг, полученных библиотекой
Института проблем управления c 20 февраля по 4 марта 2012 г.
- Актуальные проблемы радиоэлектроники и телекоммуникаций. — Самара: СГАУ, 2010. — 231 с.
- Актуальные проблемы радиоэлектроники и телекоммуникаций. Ч.2. — Самара: СГАУ, 2009. — 120 с.
- Глинченко А.С. Компьютерные спектральные измерения параметров сигналов. — Красноярск: СФУ, 2010. — 171 с.
- Информационные системы. — М.: Питер, 2011. — 539 с.
- Исследование операций. — М.: ВЦ РАН, 2010. — 90 с.
- Кулик Б.А. Алгебраический подход к интеллектуальной обработке данных и знаний. — СПб.: ПУ, 2010. — 235 с.
- Левин А.Ш.Word и Excel. — М.: Питер, 2011. — 221 с.
- Леонов В. Краткий самоучитель Windows 7. — М.: Эксмо, 2011. — 174 с.
- Леонтьев В.П. Лучшие бесплатные программы 2011. — М.: ОЛМА, 2011. — 319 с.
- Макарова Н.В. Информатика. — М.: Питер, 2011. — 573 с.
- Малинецкий Г.Г. Нелинейная динамика. — М.: URSS, 2011. — 279 c.
- Математические модели колебательных систем. – СПб.: ПУ, 2010. — 172 с.
- Меженный О.А. Microsoft Windows 7. — М.: Диалектика, 2010. — 395 с.
- Механизмы управления. — М.: URSS, 2011. — 189 c.
- Михайлов Д.В. Теоретические основы построения открытых вопросно-ответных систем. – Великий Новгород: ГУ, 2010. – 286 с.
- Молодяков С.А. Системное проектирование оптоэлектронных процессоров обработки сигналов. — СПб.: ПУ, 2011. — 226 с.
- О, Коннор Дж. Искусство системного мышления. — М.: Альпина Паблишэрз, 2011. — 250 с.
- Рогинская Л.Э. Бесконтактные и гибридные электрические аппараты высокого напряжения. — Уфа: ГАТУ, 2011. — 125 с.
- Сагалович Ю.Л. Введение в алгебраические коды. — М.: ИППИ РАН, 2010. — 302 с.
- Свиридова К. Photoshop CS4 – это просто. — СПб.: БХВ-Петербург, 2010. – 362 с.
- Сергеев А.П. Microsoft Office 2010. — М.: Диалектика, 2010. — 622 с.
- Системные исследования в энергетике. — Иркутск: ИСЭМ СО РАН, 2011. — 221 с.
- Системы проектирования технологической подготовки производства и управления этапами жизненного цикла промышленного продукта. Междунар. конф., Тезисы. — М.: ИПУ РАН, 2011. — 90 с.
- Теоретические и прикладные задачи нелинейного анализа. – М.: ВЦ РАН, 2009. — 214 с.
- Трахтенгерц Э.А. Компьютерные технологии манипулирования общественным мнением. — М.: СИНТЕГ, 2011. — 296 с.
- Тучкевич Е.И. Adobe Photoshop CS5. — СПб.: БХВ-Петербург, 2011.- 495 с.
- Уокенбах Дж. Формулы в Microsoft Excel 2010. — М.: Диалектика, 2011. — 701 с.
- Управление в сложных системах. — Уфа: УГАТУ, 2011. — 237 с.
- Федулов А.А. Введение в теорию статистически ненадёжных решений. — М.: URSS, 2011. — 277 c.
- Физика и техника высоких плотностей энергии. В.1. — Саров: ФГУП, 2011. — 333 с.
- Фробениус Ф.Г. Теория характеров и представлений групп. — М.: URSS, 2011. — 211 с.
- Харди Г.Г. Курс чистой математики. — М.: URSS, 2011. — 512 с.
- Челноков Ю.Н. Кватернионные модели и методы динамики, навигации и управления движением. — М. : Физматлит, 2011. — 556 с.
- Эванс Э. Предметно-ориентированное проектирование. — М.: Вильямс, 2011. — 443 с.
 — 231 с.
— 231 с. — М.: Диалектика, 2010. — 395 с.
— М.: Диалектика, 2010. — 395 с. Междунар. конф., Тезисы. — М.: ИПУ РАН, 2011. — 90 с.
Междунар. конф., Тезисы. — М.: ИПУ РАН, 2011. — 90 с. : Физматлит, 2011. — 556 с.
: Физматлит, 2011. — 556 с.
Список отечественных журналов
- Автоматизация и IT в энергетике. 2012. № 2.
- Автоматика и телемеханика. 2012. № 1.
- Вестник компьютерных и информационных технологий. 2012. № 2.
- Вестник Российской академии наук. 2011. Т.81. № 12; 2012. Т.82. № 1.
- Датчики и системы. 2012. № 1.
- Дифференциальные уравнения. 2011. Т. 47. № 12.
- Доклады академии наук. 2012. Т. 442. № 1, 2, 3.
- Известия ВУЗов. Проблемы энергетики. 2011. № 7-8.
- Известия СПбГЭТУ «ЛЭТИ». 2011. № 10.
- Информационные технологии. 2012. № 2 + Приложение.
- Компьютер Пресс. 2012. № 2.
- Математический сборник. 2012. Т. 203. № 1.
- Мехатроника, автоматизация, управление. 2012. № 2.
- Наука и жизнь. 2012. № 2.
- Новости космонавтики. 2012. № 2.
- Проблемы управления. 2012. № 1.
- Р.Ж. Автоматика и вычислительная техника. 2012. № 1.
- Р.Ж. Техническая кибернетика. 2012. № 1.
- Российский экономический журнал. 2011. № 6.
- Системный анализ и управление в биомедицинских системах. 2011. Т.10. № 4.
- Успехи физических наук. 2011. Т. 181. № 12.
 2012. № 2.
2012. № 2.
Список иностранных журналов
- Automation and Remote Control. 2011. V. 72. N 12; 2012. V. 73. N 1.
Картина повреждений кожи · 3dtotal · Узнать | Создать
Вступление
В этом уроке вы узнаете несколько советов и приемов, как нарисовать несколько типов повреждений кожи. Мы будем уделять особое внимание синякам, пулевым ранениям и различным типам рубцов. Я использую последнюю версию Photoshop, но вы сможете применить эти методы и техники в любом другом программном обеспечении для цифровой живописи. Также важно, чтобы у вас был какой-то планшет для рисования, я использую планшет Wacom.
Я использую последнюю версию Photoshop, но вы сможете применить эти методы и техники в любом другом программном обеспечении для цифровой живописи. Также важно, чтобы у вас был какой-то планшет для рисования, я использую планшет Wacom.
Обратите внимание, что я не использую какие-либо специальные кисти в этом уроке и что все, что здесь показано, должно быть достигнуто с помощью простого мела и аэрографа, которые должны быть доступны вам в любом программном обеспечении для цифровой живописи.
Покрасим повреждения
Синяки
Создаете ли вы текстуры для зомби или изображаете раненого персонажа, синяки полезно понимать, потому что они, скорее всего, появятся везде, где произошло повреждение кожи или плоти, что, безусловно, часто встречается в цифровом искусстве.
Первое, что следует отметить о синяках, — это то, что они бывают разных цветов. Цвет синяка указывает на его возраст и может многое рассказать вам о том, что в первую очередь послужило причиной появления синяка.
Синяк всегда бывает разных цветов
Щетка
Чтобы создать этот синяк, я использовал одну из старых кистей Photoshop Chalk с добавлением Scattering в настройках кисти.Вероятно, вы можете использовать любую текстурную кисть, чтобы добиться того же эффекта. Главное — использовать не слишком гладкую щетку, так как синяки вызваны разрывом капилляров под кожей, а поскольку между разорвавшимися капиллярами могут быть промежутки, мы хотим создать пятнистый или текстурированный вид.
Я использую кисть для мела с рассыпанием
Жизненный цикл синяка
Цвета, которые появляются в синяке, обычно обозначают различные стадии заживления и могут помочь проиллюстрировать размер и серьезность первоначальной травмы, вызвавшей синяк.После первоначальной травмы синяк обычно имеет темно-розовый цвет, так как именно здесь начинается кровотечение.![]() Обратите внимание, что интенсивность цвета будет несколько бледнеть по краям синяка по мере того, как кровотечение уменьшается по мере удаления от начальной точки травмы.
Обратите внимание, что интенсивность цвета будет несколько бледнеть по краям синяка по мере того, как кровотечение уменьшается по мере удаления от начальной точки травмы.
Со временем цвет изменится с розового на темно-фиолетовый или голубоватый (из-за гемоглобина). Во время выздоровления организм начинает разрушать гемоглобин. Разделение аминокислот и железа в гемоглобине приводит к тому, что синяк меняет цвет с пурпурного на коричнево-зеленый.На заключительном этапе процесса заживления синяк становится желтым, что вызвано образованием фермента под названием билирубин. В конечном итоге это исчезает, как только синяк полностью заживет.
Часто вы увидите синяк всех этих цветов, так как разные участки синяка могут находиться на разных этапах процесса заживления. Обычно это связано с серьезностью травмы на данном этапе. Итак, если вы можете представить, что вас ударили по руке камнем, точка контакта будет там, где произошло наибольшее повреждение, и эта точка, вероятно, пострадает больше всего, и, следовательно, заживет больше всего, в то время как внешние области будут заживать. вверх намного быстрее.Из-за этого синяк часто бывает пурпурным в центре, а затем желтоватым снаружи.
вверх намного быстрее.Из-за этого синяк часто бывает пурпурным в центре, а затем желтоватым снаружи.
Вы можете использовать это, чтобы описать тип действия, вызвавшего синяк. Например, синяк от удушения может иметь отпечаток руки фиолетового цвета вокруг шеи жертвы с более светлым синяком снаружи и так далее.
Синяки меняют цвет при заживлении
Пулевые ранения
Большинство пулевых ранений имеют как точку входа, так и точку выхода, если пуля прошла через свою жертву.Эти раны сильно отличаются друг от друга, и вы можете использовать это, чтобы помочь с повествованием, которое вы пытаетесь передать, например, если бы в вашего персонажа застрелили, когда он убегал.
Пулевые ранения будут выглядеть по-разному в зависимости от того, являются ли они входными или выходными ранениями
Щетка
Для этого я снова использовал одну из старых кистей Photoshop Chalk с добавлением Scattering в настройках. Я также использую стандартный аэрограф без дополнительных настроек.
Я также использую стандартный аэрограф без дополнительных настроек.
Я использую кисть для мела с рассыпанием и аэрограф
Пулевое входное ранение
Раны входа пули, как правило, очень чистые с минимальным кровотечением, поскольку пуля (очевидно, в зависимости от пули, но в целом) в значительной степени останавливает кровотечение, когда она проходит. Вы можете даже обнаружить, что некоторые пулевые ранения вообще не кровоточат, если пуля не пройдет сквозь тело и не покинет его. Эти раны будут иметь тенденцию быть очень темными в центре, где кровь наиболее густая и скапливается, и будут довольно светлыми по краям.Вы, вероятно, также увидите опухоль снаружи и, возможно, синяк, если рана не слишком свежая.
Выходное ранение пули
Выходные пулевые ранения, в отличие от входных, могут быть довольно грязными. Кожа и плоть будут казаться разорванными по краям, а раны могут показаться довольно серьезными. Выходные раны часто имеют большие кожные лоскуты, которые выступают из раны, откуда пуля вырвалась наружу, и они могут сопровождаться разрывами, которые могут быть маленькими или очень большими.Вы также часто будете видеть брызги крови вокруг выходной раны, через которую прошла пуля.
Выходные раны часто имеют большие кожные лоскуты, которые выступают из раны, откуда пуля вырвалась наружу, и они могут сопровождаться разрывами, которые могут быть маленькими или очень большими.Вы также часто будете видеть брызги крови вокруг выходной раны, через которую прошла пуля.
Шрамы
Существует несколько различных типов шрамов, но наиболее заметными из них являются три, которые вы, вероятно, будете использовать в рисунках и текстурах; тонкие рубцы, выпуклые рубцы, такие как келоидные и гипертрофические рубцы, а также впалые / ямчатые и атрофические рубцы. Каждый из них уникален и отличается по внешнему виду, и их можно использовать для придания большей глубины любому персонажу, потому что шрамы рассказывают историю вещей, которые происходили с нами в прошлом.
Текстура рубца
Чтобы добиться общей текстуры, имитирующей рубцовую ткань, я просто использую повторяющиеся угловые линии разных цветов.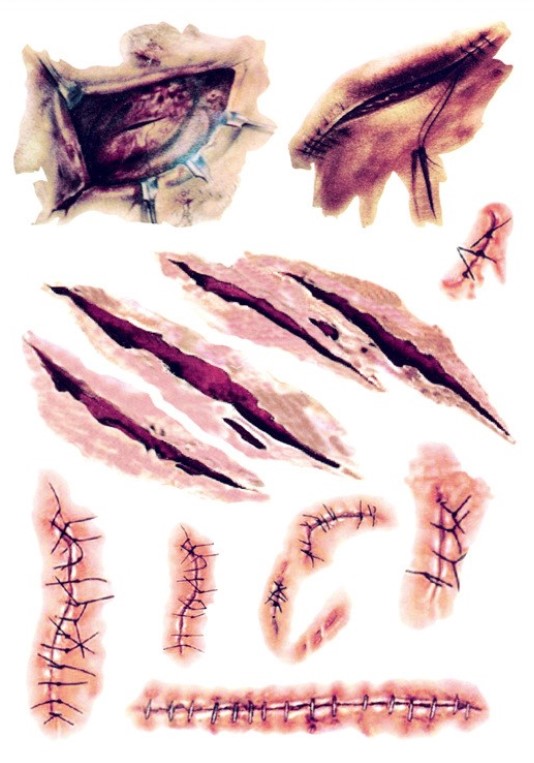 Даже зеркальные блики на шрамах используют те же угловые линии. Как и в случае с предыдущими изображениями, я использую стандартную кисть Chalk с добавлением Scattering .
Даже зеркальные блики на шрамах используют те же угловые линии. Как и в случае с предыдущими изображениями, я использую стандартную кисть Chalk с добавлением Scattering .
Используйте угловые линии для создания эффекта рубцовой ткани
Приподнятые шрамы
Рельефные рубцы выступают над кожей и возникают, когда наш организм вырабатывает слишком много коллагена в месте раны, вызывая избыток ткани.Это нарастание может продолжаться несколько месяцев, а в некоторых случаях даже лет! При гипертрофических рубцах эта ткань остается в пределах раны, но при келоидных рубцах разрастание ткани выходит за пределы границы первоначальной раны.
Тонкие рубцы
Это наиболее распространенный тип рубцов, который обычно возникает в результате обычных порезов или мелких ран. Рубцы с тонкими линиями обычно бывают плоскими и могут казаться довольно бледными или серебристыми. Поскольку эти шрамы могут быть довольно бледными, они могут быть более заметными на более темных тонах кожи, потому что ткань рубца не загорает и не темнеет со временем.При рисовании этих шрамов вы можете выбрать цвет из окружающего участка кожи и нанести некоторые из этих цветов обратно на шрам, чтобы смешать его с кожей и не допустить, чтобы он выглядел «застрявшим».
Поскольку эти шрамы могут быть довольно бледными, они могут быть более заметными на более темных тонах кожи, потому что ткань рубца не загорает и не темнеет со временем.При рисовании этих шрамов вы можете выбрать цвет из окружающего участка кожи и нанести некоторые из этих цветов обратно на шрам, чтобы смешать его с кожей и не допустить, чтобы он выглядел «застрявшим».
Впалые / ямчатые и атрофические рубцы
Эти шрамы чаще всего встречаются на лице и, как правило, возникают в результате прыщей или ветряной оспы. Обычно они появляются в виде нескольких ямок на коже, что придает коже неровную текстуру поверхности. Эти типы шрамов, как правило, не блестят, как другие, и часто даже приобретают тот же цвет, что и окружающий участок кожи.
Чтобы добиться эффекта ямок или атрофических шрамов, вы можете просто использовать немного более темный оттенок цвета кожи, чтобы создать ямки, а затем вернуть их на окружающую кожу, используя текстурную кисть с цветом окружающей кожи. Помните, что они, как правило, не блестят и не окрашены, как другие шрамы.
Помните, что они, как правило, не блестят и не окрашены, как другие шрамы.
Предыдущие уроки Джины
наложений крови, ран и шрамов [Photoshop] [8666923]
=============================================== ==========================
Загрузить наложение крови, наложения ран и шрамов [Photoshop] [8666923]
PSD
====== ================================================== =================
 Д.Все программные инструменты, руководства по графическому дизайну хорошего качества, включая фотографии в 2D / 3D анимацию, видео, доступны здесь.
Д.Все программные инструменты, руководства по графическому дизайну хорошего качества, включая фотографии в 2D / 3D анимацию, видео, доступны здесь.Те, кто являются начинающими графическими дизайнерами, хотят практиковаться с помощью обратного проектирования. Готовые материалы от Envato Elements могут сделать это, взяв Наши доступные материалы. Здесь мы сгруппировали все курсы по разным разделам, как 2D / 3D-дизайн, архитектор, объект анимированной графики, игровые графические элементы, дизайн продукта в различных программах, таких как Cinema 4D, AcceliCAD, Adobe Animate, After Effects, ArchiCAD, CAD Modeling, 3ds. Max, Autocad, AutoDesk, Blender, Catia, ColorWay, MARI, Maya, MODO, MotionBuilder, Mudbox, NUKE, Revit, Sketchbook, Sketchup, Softimage, Solid Works, V-Ray, ZBrush, Camtasia, Character Animator, Цифровая живопись, Final Cut Pro, Unity, Rhino, Terragen и многое другое.
Наложение Хэллоуина и цифровой фон Хэллоуина: Наложение Photoshop, наложение с привидениями, наложение ведьмы, тыква png, горшок png, череп клипарт
✰Подробнее: 24 Хэллоуин оверлей / png / клипарт 300 dpi высокое разрешение
Ваши фотографии на Хэллоуин скучные, и вы хотите добавить в них немного волшебства? с нашими изображениями вы создадите идеальный дизайн.
Этот оверлей для фотошопа: содержит 24 уникальных слоя для Хэллоуина. Получайте великолепные потрясающие изображения от профессионалов фотографии и используйте их в качестве эффекта, наложения, элемента изображения или фона.
Просто выберите эффекты с помощью Photoshop на своем изображении — вот и все!
Совместим с любым программным обеспечением, позволяющим работать со слоями (Photoshop, Photoshop Elements, Paint Shop Pro, PicMonkey’s Photo Editor и т. Д.).
Примечание для пользователей Lightroom — убедитесь, что у вас есть плагин, позволяющий работать с слои
Загрузить наложение крови, наложения ран и шрамов [Photoshop] [8666923] Бесплатно
Предварительный просмотр проекта:
🌟 Загрузить еще бесплатный контент 🌟
✪ Загрузить шаблоны After Effect ✪ Загрузить шаблоны Premiere Pro ✪ Загрузить бесплатный курс ✪ Бесплатные проекты Videohive ✪ Бесплатные векторные изображения ✪ Загрузить переходы ✪ Бесплатно Photoshop PSD ✪ Шаблон баннера и макета ✪ Загрузить текстуры и узоры ✪
Бесплатное обучение графическому дизайну для создания красивого графического дизайна
Зарегистрируйте учетную запись Pro с помощью ClicknUpload | FileUpload для быстрой загрузки
Графические знаки | Xtreme Graphix
Скачать бесплатные быстрые ссылки
Без премиальных ссылок, только бесплатные быстрые ссылки
Пожалуйста, потратьте 30 секунд и оставьте комментарий | Нравится | Поделиться | Следуйте | Подписаться
============
Как скачать
============
Загрузить | Zippyshare
Пароль: (если требуется) [GraphixTree. com]
com]
:: ТОЛЬКО ДЛЯ ОЦЕНКИ И ОБРАЗОВАТЕЛЬНЫХ ЦЕЛЕЙ ::
:: Перед загрузкой этого выпуска щелкните здесь и прочтите! ::
Примечание:
Если есть неработающая ссылка, оставьте комментарий ниже.
Dead Link будет повторно загружен в течение 1-24 часов.
Теги:
скачать бесплатные макеты psd | скачать бесплатные шаблоны psd | скачать бесплатные psd файлы | скачать бесплатно PSD баннер | скачать бесплатно psd кисти | графический дизайн | графические знаки | курс графического дизайна | Бесплатные шаблоны Photoshop | Бесплатные шаблоны After Effects | Скачать бесплатно EPS | Скачать бесплатные шаблоны VideoHive
(Посещали 92 раза, сегодня 1 раз)
Как сделать раны в Photoshop
Реалистичная рана и кровь в Photoshop. Реалистичная рана и кровь в фотошопе. Джессика Рэнделл 24 октября 2013 г. Эффект рисования, Учебное пособие по Photoshop. Существует не так много руководств о нанесении ран или крови. Здесь вы можете попробовать. Это особенно полезно при создании сцены боя. Создайте рану в фотошопе. Добавить в новый плейлист. Вы уже сообщили об этом видео. Мы ценим вашу помощь. Вы уже проголосовали за этот ролик. Вот что происходит, когда корова влюбляется.Только я имею на это право, человек! У тебя красные булочки, хун! 24 октября 2013 г. — Существует не так много руководств о нанесении ран или крови. Здесь вы можете иметь файл. Советы по созданию фотографий 12 июня 2019 г.
Существует не так много руководств о нанесении ран или крови. Здесь вы можете попробовать. Это особенно полезно при создании сцены боя. Создайте рану в фотошопе. Добавить в новый плейлист. Вы уже сообщили об этом видео. Мы ценим вашу помощь. Вы уже проголосовали за этот ролик. Вот что происходит, когда корова влюбляется.Только я имею на это право, человек! У тебя красные булочки, хун! 24 октября 2013 г. — Существует не так много руководств о нанесении ран или крови. Здесь вы можете иметь файл. Советы по созданию фотографий 12 июня 2019 г.
- Как сделать раны в Photoshop Видео
- Как сделать раны в фоновом режиме Photoshop
- Как сделать раны в Photoshop Скачать
Инструмент выбора Magic Wand был с тех пор, как Photoshop был в зачаточном состоянии, а Elements еще не была мерцанием в глазах Adobe.Его чрезвычайно легко использовать, но немного сложнее предсказать, какие результаты выбора он представит.
Вот как это работает: вы щелкаете внутри изображения, и инструмент «Волшебная палочка» делает выбор.![]() Этот выбор основан на цвете пикселя, по которому вы щелкнули. Если другие пиксели похожи по цвету на ваш целевой пиксель, Elements включает их в выбор. Параметр Tolerancesetting определяет , насколько должен быть похожий цвет, чтобы инструмент Magic Wand выбрал его.
Этот выбор основан на цвете пикселя, по которому вы щелкнули. Если другие пиксели похожи по цвету на ваш целевой пиксель, Elements включает их в выбор. Параметр Tolerancesetting определяет , насколько должен быть похожий цвет, чтобы инструмент Magic Wand выбрал его.
Говоря о допуске
Параметр «Допуск» определяет диапазон цветов, выбираемых инструментом «Волшебная палочка».Он основан на уровнях яркости от 0 до 255:
При установке значения «Допуск» на 0 выбирается только один цвет.
При установке параметра «Допуск 255» выбираются все цвета или все изображение.
Значение по умолчанию — 32, поэтому всякий раз, когда вы щелкаете пиксель, Elements анализирует значение этого основного цвета, а затем выбирает все пиксели, уровни яркости которых находятся между 16 уровнями светлее и 16 уровнями темнее.
Что делать, если изображение содержит несколько оттенков одного цвета? Это не большая проблема.Вы можете сделать несколько щелчков Волшебной палочки, чтобы выбрать дополнительные пиксели, которые вы хотите включить в выделение. Или вы можете попробовать установить более высокий допуск. И наоборот, если ваша палочка выбирает слишком много, вы также можете снизить настройку допуска.
Или вы можете попробовать установить более высокий допуск. И наоборот, если ваша палочка выбирает слишком много, вы также можете снизить настройку допуска.
Инструмент «Волшебная палочка» лучше всего работает, когда у вас есть высококонтрастные изображения или изображения с ограниченным количеством цветов. Не используйте инструмент «Волшебная палочка», если в изображении много цветов и нет реального четкого контраста между желаемым элементом и фоном.
Использование палочки для выбора допуска
Чтобы использовать инструмент «Волшебная палочка» для настройки допуска, выполните следующие действия:
Выберите инструмент «Волшебная палочка» на панели «Инструменты».
Похоже на жезл со звездочкой на конце. Вы также можете просто нажать W.
Щелкните в любом месте на желаемом элементе, используя значение Допуска по умолчанию 32.
Помните, что пиксель, который вы щелкаете, определяет базовый цвет.
Если вы выбрали все, что хотите, при первом нажатии, все готово.
Если ваш выбор требует дальнейшей настройки, как верхнее изображение на следующем рисунке, перейдите к шагу 3.Magic Wand выбирает пиксели на основе заданной настройки допуска.
Укажите новую настройку допуска на панели параметров.
Если Magic Wand выбирает больше, чем вы хотите, уменьшите настройку Tolerance. Если жезл выбрал недостаточно, увеличьте значение.
Пока вы осматриваете панель параметров, вот еще пара параметров, с которыми вам нужно ознакомиться:
Anti- A lias: Смягчает край выделения на один ряд пикселей.
Смежные: Заставляет Magic Wand выбирать только пиксели, которые находятся рядом друг с другом. Без этой опции инструмент выбирает все пиксели в пределах допуска, независимо от того, смежны они друг с другом или нет.
Пример Все слои: Если у вас несколько слоев и включена эта опция, Magic Wand выбирает пиксели из всех видимых слоев. Без этой опции инструмент выбирает пиксели только из активного слоя.
Уточнить края: Очистите выделение, переместив ползунок «Сглаживание», чтобы уменьшить количество неровностей на краях. Переместите ползунок Contract / Expand влево или вправо, чтобы уменьшить или увеличить выбранную область соответственно. Нажмите кнопку настраиваемого цвета наложения, чтобы просмотреть свой выбор со скрытыми краями и наложением цвета только экрана в невыделенной области.
Щелкните нужный элемент еще раз.
К сожалению, волшебная палочка недостаточно волшебна, чтобы автоматически изменить ваш первый выбор.Вместо этого он отменяет выделение текущего выбора и делает новый выбор на основе вашей новой настройки допуска. Если по-прежнему что-то не так, вы можете снова настроить параметр «Допуск».
 Если ваш выбор требует дальнейшей настройки, как верхнее изображение на следующем рисунке, перейдите к шагу 3.
Если ваш выбор требует дальнейшей настройки, как верхнее изображение на следующем рисунке, перейдите к шагу 3.Как сделать раны в Photoshop Видео
Люди используют искусство макияжа для создания реалистичных физических деталей для визуального эффекта. Вы можете сделать великолепно выглядящие, реалистичные раны для фильмов, костюмированных вечеринок или чего угодно, что только можно придумать. Вы можете создать все, что угодно. Все, что вам нужно, это несколько припасов, отличный вкус и ваше воображение.Вот как можно создать искусственные шрамы и раны.
Порядок действий:
Вещи, которые вам понадобятся;
- Смесь желатина без вкусовых добавок
- 1/4 стакана кипятка
- Медицинский спирт
- Ватные шарики
- Неиспользованные палочки от эскимо
- Вазелин
- Тональный крем для макияжа
- Темно-красный румянец
- Аппликатор теней
- Ватные палочки
Шаг 1
Вы всегда хотите начинать с чистой палитры.Уберите волосы с лица. Закрепите волосы заколкой или галстуком. Тщательно вымойте лицо мягким очищающим средством и водой. Промокните кожу насухо.
Шаг 2
Смочите спиртом ватный диск. Вытрите то место, где хотите нанести фальшивку. Например, лоб. Это помогает макияжу держаться на коже. Избыток спирта быстро испарится.
Шаг 3
Кипятите ¼ стакана воды. Добавьте в воду неароматизированную желатиновую смесь.Смешайте желатин и воду с помощью палочки для мороженого. Дайте желатину остыть и загустеть. Желатин должен быть горячим, но безопасным для нанесения на кожу. Кончиками пальцев проверяйте температуру желатина, пока он остывает. Будьте осторожны.
Как сделать раны в фоновом режиме Photoshop
Шаг 4
Нанесите немного вазелина на ватный тампон. Втирайте брови вазелином. Это защитит волосяные фолликулы бровей от приклеивания к горячему желатину.Этот шаг важен. Когда вы будете готовы удалить искусственную рану, это поможет.
Шаг 5
Используйте хорошо отведенное рабочее пространство. Слегка наклоните голову назад и под углом, чтобы вы могли четко видеть, как продвигается ваша работа. Нанесите на лоб небольшую чайную ложку желатина. Сделайте это так, чтобы желатин стекал с аппликатора на поверхность вашей кожи. Вы должны убедиться, что желатин падает по диагонали. Прежде чем двигаться дальше, дайте желатину застыть.
Step 6
Как сделать раны в Photoshop Скачать
Примените еще одну чайную ложку желатина ко лбу, используя тот же метод. На этот раз позвольте желатину немного упасть выше последнего нанесения. Это создаст отдельные складки с промежутком между ними. Дайте им время застыть. Когда складки затвердеют, нанесите на раны немного жидкого тонального крема. Это придает им реалистичный вид кожи. С помощью другого хлопкового лебедя нанесите темно-красный румянец на внутреннюю часть разреза.Это та часть, которая придает ране вид срезанной кожи. Или вы всегда можете использовать поддельную кровь. Это легко найти и применить.
Scar Booth в App Store
Подделайте своих друзей, добавляя реалистичные шрамы, порезы и синяки на свои фотографии с помощью приложения для iPhone от Scar Booth. Сделайте любой снимок и добавьте на фото виртуальные раны.
Как видно из телешоу «Доктора» и раскритиковано Daily Mail за то, что он слишком реалистичен.
Отрегулируйте прозрачность и размытость изображения, чтобы оно выглядело так, как будто это настоящая травма.
Ваши друзья и семья будут уверены, что вы получили болезненную травму, когда увидят вашу фотографию с синяком под глазом, синяками и изрезанным лицом. Между тем, вы выглядите как никогда хорошо и только что опубликовали шутливую фотографию.
Представьте себе комментарии, которые вы получите, когда разместите на Facebook свою фотографию со шрамами, порезами и синяками, и никто не знает, что за этим стоит.
Отличные отзывы пользователей
«Dr — Хорошее приложение для развлечения, и мои дети тоже думают, что оно хорошо»
«Великолепно — очень нравится»
«Так весело — классное приложение»
«Koolness — Totally Koop luv it! Это супер страшно! Я зомбировал свою маму, себя и сестру! Я полностью обожаю это приложение из сети! »
«Самое забавное приложение на свете — я так повеселился, изувечив себя и остальных членов своей семьи.Я отправил их в fb и обманул всех »
Features
— Универсальное приложение для iPhone и iPad
— Retina Graphics
— Более 100 уникальных изображений травм и их рост
— Сделайте снимок или загрузите один из альбома
— Поделиться через всех популярные социальные платформы
Расширенное редактирование
— Масштабирование двойным касанием для увеличения и внесения мелких изменений
— Регулировка уровня прозрачности травм
— Ластик, стирание частей раны
— Размытие, регулировка размытия раны
— Яркость, отрегулируйте яркость раны
— Добавьте несколько травм к фотографии
— Отрегулируйте размер, поворот и расположение
Загрузите Scar Booth сейчас и начните делать шалости с фотографиями травм
Есть твиттер? не поощряйте насилие по отношению к кому-либо или чему-либо! Это приложение было создано как забавное приложение для розыгрыша и не более того.
Сравнение трех методов измерения площади раны: Photoshop, ImageJ и Visitrak
1. Haghpanah S, Bogie K, Wang X, Banks PG, Ho CH. Надежность электронного измерения по сравнению с ручным измерением ранытехника. Arch Phys Med Rehabil. 2006; 87: 1396-1402.
2. Sheehan P, Jones P, Caselli A, Giurini J, Veves A. Процентное изменение площади раны язв диабетической стопы за 4-недельный период является надежным предиктором полного заживления в 12-недельном проспективном исследовании. Уход за диабетом.2003; 26 (6): 1879-
.
82.
3. Hammond CE. ARANZ Medical SilhouetteTM: инновационный прибор для измерения и документирования ран
система. Перспективы неотложной помощи. 2008; Летом: 12-5.
4. Литтл К., Макдональд Дж., Дженкинс М.Г., Маккоррон П. Обзор методов, используемых для измерения площади раны и
объем. J Уход за раной. 2009; 18 (16): 250-3.
5. Шоу Дж., Хьюз К.М., Лаган К.М. Оценка трех методов измерения ран при диабетической стопе.
Уход за диабетом. 2007; 30 (10): 2641-2.
6. Чанг А.С., Дирман Б., Гринвуд Дж. Э. Сравнение методов измерения площади раны: Visitrak и
фотография. EPlasty. 2011; 11: 158-66.
7. Li PN, Li H, Wu ML, Wang SY, Kong QY, Zhen Z. et al. Экономичное цифровое изображение на основе прозрачности
для эффективного и точного измерения площади раны. PLoS One. 2012; 7 (5): e38069. DOI: 10.1371 / journal.pone.
0038069
8. Джунорн Т., Каногсунторнтрат Н, Оратхай П.Обоснованность методик измерения площади раневой поверхности. Рама Нурс Дж.
2014; 20 (3): 314-23. (на тайском языке)
9. Sugama J, Matsui Y, Sanada H, Konya C, Okuwa M, Kitagawa A. Исследование эффективности и удобства
передовая портативная система измерения ран (VisitrakTM). J Clin Nurs. 2007; 16: 1265-9.
10. Фаул Ф., Эрдфельдер Э., Ланг А. Г., Бюхнер А. Power 3: гибкая программа статистического анализа власти для социальных сетей,
поведенческие и биомедицинские науки.Методы Behav Res. 2007; 39 (2): 175-91.
11. van Zuijlen, PPM, Angeles AP, Suijker MHK Reis RW, Middelkoop E. Надежность и точность практического применения
методы измерения площади поверхности ран и шрамов. Int J Low Extrem Wounds, 2004; 3 (1): 7–11.
12. Портни Л.Г., Уоткинс М.П. Основы клинических исследований: приложения к практике. 3-е изд. Верхняя река Сэдл:
Пирсон / Прентис Холл; 2015.
13. Харрис-Лав МО, Симон Б.А., Тейшейра К., Исмаил К.Ультразвуковые оценки качества мышц у пожилых людей:
надежность и сравнение Photoshop и Image J для анализа мышечной эхогенности в оттенках серого. Пир Дж.
2016; 4: e1721. DOI: 10.7717 / peerj.1721.
14. Стоктон К.А., Макмиллиан К.М., Стори К.Дж., Дэвид М.К., Кимбл Р.М. 3D-фотография так же точна, как цифровая
рентгеноскопия при определении площади ожоговой раны. Бернс, 2015; 41: 80-4.
15. Бхеди А., Саксена А. К., Гадани Р., Патель Р. Цифровая фотография и методы на основе прозрачности для
измерение площади раневой поверхности.Индийский журнал J Surg.2013; 75 (2): 111–14. DOI: 10.1007 / s12262-012-0422-y
16. Тан XN, Берман AE, Swanson RA, Yenari MA. Цифровое количественное определение кровоизлияния в мозг с помощью Photoshop и
Изображение J. J. Neurosci. Методы. 2010. 190 (2): 240–3.
Hedgehog стимулирует регенерацию волосяных фолликулов, создавая индуктивную дерму во время заживления ран на коже мыши.
Sennett, R. & Rendl, M. Мезенхимно-эпителиальные взаимодействия во время морфогенеза и цикличности волосяного фолликула. Семин. Cell Dev. Биол. 23 , 917–927 (2012).
CAS Статья Google ученый
Харди М. Х. Тайная жизнь волосяного фолликула. Trends Genet. 8 , 55–61 (1992).
CAS Статья Google ученый
Морган, Б.А. Дермальный сосочек: поучительная ниша для эпителиальных стволовых клеток и клеток-предшественников в развитии и регенерации волосяного фолликула. Колд Спринг Харб. Перспектива. Med. 4 , a01518 (2014).
Артикул Google ученый
Rompolas, P. & Greco, V. Динамика стволовых клеток в нише волосяного фолликула. Семин. Cell Dev. Биол. 25–26 , 34–42 (2014).
Артикул Google ученый
Cotsarelis, G., Sun, T. & Lavker, R.M. Клетки, сохраняющие метку, находятся в области выпуклости волосяного покрова: влияние на фолликулярные стволовые клетки, цикл роста волос и канцерогенез кожи. Cell 61 , 1329–1337 (1990).
CAS Статья Google ученый
Blanpain, C. & Fuchs, E. Эпидермальный гомеостаз: балансирующее действие стволовых клеток в коже. Нац. Rev. Mol. Cell Biol. 10 , 207–217 (2009).
CAS Статья Google ученый
Шнайдер М. Р., Шмидт-Ульрих Р. и Паус Р.Волосяной фолликул как динамический миниорган. Curr. Биол. 19 , R132 – R142 (2009 г.).
CAS Статья Google ученый
Gurtner, G.C., Werner, S., Barrandon, Y. & Longaker, M.T. Ремонт и регенерация ран. Природа 453 , 314–321 (2008).
ADS CAS Статья Google ученый
Зейферт, А.W. & Maden, M. Новые взгляды на регенерацию кожи позвоночных. Внутр. Rev. Cell Mol. Биол. 310 , 129–169 (2014).
CAS Статья Google ученый
Leavitt, T. et al. Заживление ран без рубцов: поиск нужных клеток и сигналов. Cell Tissue Res. 365 , 483–493 (2016).
CAS Статья Google ученый
Ito, M. et al. Wnt-зависимая регенерация волосяных фолликулов de novo в коже взрослых мышей после ранения. Природа 447 , 316–320 (2007).
ADS CAS Статья Google ученый
Чуонг, К. М. Регенеративная биология: новые волосы из заживляющих ран. Природа 447 , 265–266 (2007).
ADS CAS Статья Google ученый
Straile, W. E. in Advances in Biology of Skin , Vol. 9 (ред. Монтанья, В. и Добсон, Р.) 369–391 (Pergamon Press, Нью-Йорк, 1967).
Chiang, C. et al. Существенная роль Sonic hedgehog в морфогенезе волосяного фолликула. Dev. Биол. 205 , 1–9 (1999).
CAS Статья Google ученый
Гали, Л., Вонг, С. Т., Грин, Дж., Тидман, Н. и Куинн, А.Белок G. Gli1 экспрессируется в базальноклеточных карциномах, кератиноцитах наружной корневой оболочки и в субпопуляции мезенхимальных клеток нормальной кожи человека. J. Invest. Дерматол. 113 , 595–599 (1999).
CAS Статья Google ученый
St-Jacques, B. et al. Звуковой сигнал ежа важен для развития волос. Curr. Биол. 8 , 1058–1068 (1998).
CAS Статья Google ученый
Woo, W. M., Zhen, H. H. & Oro, A. E. Shh поддерживает идентичность дермальных сосочков и морфогенез волос посредством регуляторной петли Noggin-Shh. Genes Dev. 26 , 1235–1246 (2012).
CAS Статья Google ученый
Успенская Т., Матос И., Мерц А. Ф., Фиоре В. Ф. и Фукс Е. Антагонизм WNT-SHH определяет и расширяет стволовые клетки до образования ниши. Ячейка 164 , 156–169 (2016).
CAS Статья Google ученый
Gao, J. et al. Ламинин-511 — это эпителиальный сигнал, способствующий развитию и функционированию дермальных сосочков во время раннего морфогенеза волос. Genes Dev. 22 , 2111–2124 (2008).
CAS Статья Google ученый
Сато Н., Леопольд П. Л. и Кристал Р. Г. Индукция фазы роста волос у постнатальных мышей путем локальной временной экспрессии Sonic hedgehog. J. Clin. Инвестировать. 104 , 855–864 (1999).
CAS Статья Google ученый
Сато Н., Леопольд П. Л. и Кристал Р. Г. Влияние опосредованной аденовирусом экспрессии гена Sonic hedgehog на отрастание волос у мышей с алопецией, вызванной химиотерапией. J. Natl. Cancer Inst. 93 , 1858–1864 (2001).
CAS Статья Google ученый
Морган, Б.А., Оркин, Р.В., Норамли, С. и Перес, А. Стадийные эффекты экспрессии звукового ежа в эпидермисе. Dev. Биол. 201 , 1–12 (1998).
CAS Статья Google ученый
Wang, D. H. et al. Аберрантная эпителиально-мезенхимальная передача сигналов Hedgehog характеризует метаплазию Барретта. Гастроэнтерология 138 , 1810–1822 (2010).
CAS Статья Google ученый
Zhou, P., Byrne, C., Jacobs, J. & Fuchs, E. Фактор 1 лимфоидного энхансера управляет формированием рисунка волосяного фолликула и судьбой эпителиальных клеток. Genes Dev. 9 , 700–713 (1995).
CAS Статья Google ученый
Бочкарев В.А. и др. Noggin — это мезенхимный стимулятор индукции волосяных фолликулов. Нац. Cell Biol. 1 , 158–164 (1999).
CAS Статья Google ученый
Sennett, R. et al. Интегрированный транскриптомный атлас предшественников эмбриональных волосяных фолликулов, их ниш и развивающейся кожи. Dev. Клетка. 34 , 577–591 (2015).
CAS Статья Google ученый
Меркель, Дж. Р., ДиПаоло, Б. Р., Хэллок, Г. Г. и Райс, Д. С. Содержание коллагена типа I и типа III в заживающих ранах у плодов и взрослых крыс. Proc. Soc. Exp. Биол. Med. 187 , 493–497 (1988).
CAS Статья Google ученый
Whitby, D. J. & Ferguson, M. W. Внеклеточный матрикс ран губ у эмбрионов, новорожденных и взрослых мышей. Развитие 112 , 651–668 (1991).
CAS PubMed Google ученый
Коллинз, К. А., Кречмар, К. и Ватт, Ф. М. Перепрограммирование дермы взрослого человека в неонатальное состояние посредством эпидермальной активации бета-катенина. Развитие 138 , 5189–5199 (2011).
CAS Статья Google ученый
Seifert, A. W. et al. Отшелушивание кожи и регенерация тканей у африканских колючих мышей (Acomys). Природа 489 , 561–565 (2012).
ADS CAS Статья Google ученый
Дуайон, К. Дж., Данн, М. Г. и Сильвер, Ф.З. Взаимосвязь механических свойств и структуры коллагена закрытых и открытых ран. J. Biomech. Англ. 110 , 352–356 (1988).
CAS Статья Google ученый
West, J. et al. Легочная гипертензия у трансгенных мышей, экспрессирующих доминантно-отрицательный ген BMPRII в гладких мышцах. Circ. Res. 94 , 1109–1114 (2004).
CAS Статья Google ученый
Wixler, V. et al. Дефицит белка Fhl2, состоящего только из LIM, ухудшает заживление кожных ран. J. Cell Biol. 177 , 163–172 (2007).
CAS Статья Google ученый
Plikus, M. V. et al. Регенерация жировых клеток из миофибробластов при заживлении ран. Наука 355 , 748–752 (2017).
ADS CAS Статья Google ученый
Zhang, Y. et al. Активация передачи сигналов бета-катенина программирует эмбриональный эпидермис на судьбу волосяного фолликула. Развитие 135 , 2161–2172 (2008).
CAS Статья Google ученый
Lynch, MH, O’Guin, WM, Hardy, C., Mak, L. & Sun, TT Кислые и основные кератины волос / ногтей («твердые»): их колокализация в верхних кортикальных клетках и клетках кутикулы волосяного фолликула человека и их отношение к «мягким» кератинам. J. Cell Biol. 103 , 2593–2606 (1986).
CAS Статья Google ученый
О’Гин, В. М., Сан, Т. и Манабе, М. Взаимодействие трихогиалина с промежуточными филаментами: три иммунологически определенные стадии созревания трихогиалина. J. Invest. Дерматол. 98 , 24–32 (1992).
Артикул Google ученый
Лю Ю., Лайл С., Янг З. и Котсарелис Г. Промотор кератина 15 нацелен на предполагаемые эпителиальные стволовые клетки в выпуклости волосяного фолликула. J. Invest. Дерматол. 121 , 963–968 (2003).
CAS Статья Google ученый
Trempus, C. S. et al. Обогащение живых кератиноцитов мышей из выпуклости волосяного фолликула маркером клеточной поверхности CD34. J. Invest. Дерматол. 120 , 501–511 (2003).
CAS PubMed Google ученый
Jahoda, C. A., Reynolds, A. J., Chaponnier, C., Forester, J. C. & Gabbiani, G. Альфа-актин гладких мышц является маркером дермы волосяного фолликула in vivo и in vitro. J. Cell Sci. 99 (Pt 3), 627–636 (1991).
PubMed Google ученый
Lawson, D., Harrison, M. & Shapland, C.Трансгелин фибробластов и SM22alpha гладких мышц — это один и тот же белок, экспрессия которого подавляется во многих клеточных линиях. Cell Motil. Цитоскелет. 38 , 250–257 (1997).
CAS Статья Google ученый
Grieskamp, T., Rudat, C., Ludtke, T.H., Norden, J. & Kispert, A. Передача сигналов Notch регулирует дифференцировку гладких мышц клеток, происходящих из эпикарда. Circ. Res. 108 , 813–823 (2011).
CAS Статья Google ученый
Clements, M. P. et al. Микроокружение раны перепрограммирует шванновские клетки в инвазивные мезенхимальные клетки, чтобы управлять регенерацией периферических нервов. Neuron 96 , 98–114.e117 (2017).
CAS Статья Google ученый
Ellinghaus, P. et al. Экспрессия провоспалительных генов в эндотелиальных клетках человека: сравнение ривароксабана и дабигатрана. Тромб. Res. 142 , 44–51 (2016).
CAS Статья Google ученый
Rezza, A. et al. Сигнальные сети среди предшественников стволовых клеток, предшественников, усиливающих транзит, и их ниши в развивающихся волосяных фолликулах. Cell Rep. 14 , 3001–3018 (2016).
CAS Статья Google ученый
Рендл, М., Льюис, Л. и Фукс, Э. Молекулярное рассечение мезенхимно-эпителиальных взаимодействий в волосяном фолликуле. PLoS Biol. 3 , e331 (2005).
Артикул Google ученый
Дрискелл, Р. Р., Клавель, К., Рендл, М. и Ватт, Ф. М. Клетки дермального сосочка волосяного фолликула: краткий обзор. J. Cell Sci. 124 , 1179–1182 (2011).
CAS Статья Google ученый
Rendl, M., Polak, L. & Fuchs, E. Передача сигналов BMP в клетках дермального сосочка необходима для их индуктивных свойств волосяного фолликула. Genes Dev. 22 , 543–557 (2008).
CAS Статья Google ученый
Xie, J. et al. Активация сглаженных мутаций при спорадической базальноклеточной карциноме. Nature 391 , 90–92 (1998).
ADS CAS Статья Google ученый
Wong, S. Y. & Reiter, J. F. Ранение мобилизует стволовые клетки волосяного фолликула для образования опухолей. Proc. Natl Acad. Sci. США 108 , 4093–4098 (2011).
ADS CAS Статья Google ученый
Youssef, K. K. et al. Идентификация клеточной линии происхождения базальноклеточной карциномы. Нац. Cell Biol. 12 , 299–305 (2010).
CAS Статья Google ученый
Yang, S.H. et al. Патологические ответы на онкогенную передачу сигналов Hedgehog в коже зависят от канонической передачи сигналов Wnt / beta3-catenin. Нац. Genet. 40 , 1130–1135 (2008).
CAS Статья Google ученый
Gay, D. et al. Fgf9 из дермальных гаммадельта-Т-клеток индуцирует регенерацию волосяных фолликулов после ранения. Нац. Med. 19 , 916–923 (2013).
CAS Статья Google ученый
Chen, D., Jarrell, A., Guo, C., Lang, R. & Atit, R. Активность дермального бета-катенина в ответ на эпидермальные лиганды Wnt необходима для пролиферации фибробластов и инициации волосяных фолликулов. Разработка 139 , 1522–1533 (2012).
CAS Статья Google ученый
Wang, X. et al. Макрофаги индуцируют AKT / бета-катенин-зависимую активацию Lgr5 (+) стволовых клеток и регенерацию волосяных фолликулов через TNF. Нац. Commun. 8 , 14091 (2017).
ADS CAS Статья Google ученый
Nelson, A. M. et al. дцРНК, высвобождаемая при повреждении тканей, активирует TLR3 для ускорения регенерации кожи. Стволовые клетки клеток 17 , 139–151 (2015).
CAS Статья Google ученый
Hamburg-Shields, E., DiNuoscio, G.J., Mullin, N.K., Lafyatis, R. & Atit, R.P. Устойчивая активность бета-катенина в дермальных фибробластах способствует фиброзу за счет активации экспрессии генов, кодирующих белок внеклеточного матрикса. J. Pathol. 235 , 686–697 (2015).
CAS Статья Google ученый
Rognoni, E. et al. Ингибирование передачи сигналов бета-катенина в дермальных фибробластах усиливает регенерацию волосяных фолликулов во время заживления ран. Разработка 143 , 2522–2535 (2016).
CAS Статья Google ученый
Choi, Y. S. et al. Определенные функции Wnt / бета-катенина в пролиферации и выживании стволовых клеток волосяных фолликулов и в межфолликулярном эпидермальном гомеостазе. Стволовые клетки клеток 13 , 720–733 (2013).
CAS Статья Google ученый
Ferrer-Vaquer, A. et al. Чувствительный и яркий одноклеточный репортер передачи сигналов Wnt / ss-catenin в реальном времени. BMC Dev. Биол. 10 , 121 (2010).
CAS Статья Google ученый
Gat, U., DasGupta, R., Degenstein, L. & Fuchs, E. Морфогенез волосяного фолликула De novo и опухоли волос у мышей, экспрессирующих усеченный бета-катенин в коже. Cell 95 , 605–614 (1998).
CAS Статья Google ученый
Oro, A. E. et al. Базальноклеточный рак у мышей со сверхэкспрессией sonic hedgehog. Наука 276 , 817–821 (1997).
CAS Статья Google ученый
Такео, М., Ли, В. и Ито, М. Заживление ран и регенерация кожи. Колд Спринг Харб. Перспектива. Med. 5 , a023267 (2015).
Артикул Google ученый
Myung, P. S., Takeo, M., Ito, M. & Atit, R. P. Секреция эпителиального лиганда Wnt необходима для роста и регенерации волосяного фолликула взрослого человека. J. Invest. Дерматол. 133 , 31–41 (2013).
CAS Статья Google ученый
Harada, N. et al. Полипоз кишечника у мышей с доминантной стабильной мутацией гена бета-катенина. EMBO J. 18 , 5931–5942 (1999).
CAS Статья Google ученый
Rabbani, P. et al. Скоординированная активация Wnt в эпителиальных и стволовых клетках меланоцитов инициирует регенерацию пигментированных волос. Cell 145 , 941–955 (2011).
CAS Статья Google ученый
Ito, M. et al. Стволовые клетки в выпуклости волосяного фолликула способствуют заживлению ран, но не способствуют. Нац. Med. 11 , 1351–1354 (2005).
CAS Статья Google ученый
Takeo, M. et al. EdnrB управляет регенеративным ответом стволовых клеток меланоцитов за счет перекрестных помех с передачей сигналов Wnt. Cell Rep. 15 , 1291–1302 (2016).
CAS Статья Google ученый
Хуанг да, В., Шерман, Б. Т. и Лемпицки, Р.A. Систематический и комплексный анализ больших списков генов с использованием ресурсов биоинформатики DAVID. Нац. Protoc. 4 , 44–57 (2009).
Артикул Google ученый
Takeo, M. et al. Активация Wnt в эпителии ногтя связывает рост ногтя с регенерацией пальца. Природа 499 , 228–232 (2013).
ADS CAS Статья Google ученый
Нарушение заживления ран с дефектной экспрессией хемокинов и привлечением миелоидных клеток у мышей с дефицитом TLR3
Abstract
Повреждение кожи вызывает как врожденный, так и адаптивный иммунный ответ для восстановления целостности ткани.TLR играют решающую роль в ответах хозяина на оскорбления. Предыдущие исследования показали, что РНК, высвобождаемые из поврежденных тканей, служат эндогенными лигандами для TLR3. В этом исследовании мы исследовали участие TLR3 в восстановлении кожи после травмы. На коже мышей с дефицитом TLR3 были созданы полные иссечные раны. Мы обнаружили, что закрытие кожной раны у мышей TLR3 — / — происходило значительно позже, чем у контрольных однопометников. Параметры заживления ран, включая реэпителизацию, образование грануляций и неоваскуляризацию, были снижены у мышей TLR3 — / — .Дальнейшие исследования показали, что отсутствие TLR3 приводит к неправильному рекрутированию нейтрофилов и макрофагов в сочетании со сниженной экспрессией хемокинов, MIP-2 / CXCL2, MIP-1α / CCL3 и MCP-1 / CCL2 в ране. Более того, у мышей дикого типа уровень мРНК и содержание белка TLR3 были значительно повышены в раненой коже и подавление адаптера сигнала TLR3, содержащего домен Toll / IL-1R, индуцирующего IFN-β с малой интерферирующей РНК, замедляющей закрытие ран. Эти результаты указывают на важную роль адаптера, содержащего домен TLR3 и Toll / IL-1R, в индукции IFN-β в заживлении ран путем регулирования продукции хемокинов и привлечения миелоидных клеток в рану для восстановления тканей.
Повреждение кожи активирует как врожденный, так и адаптивный иммунный ответ хозяина для восстановления целостности ткани (1). Заживление ран, инициированное травмой, микробами или инородными материалами, — это хорошо скоординированный процесс. Весь процесс восстановления можно разделить на четыре отдельных, но частично совпадающих фазы: немедленный ответ, воспаление, регенерация и ремоделирование (2). В большинстве органов повреждение затрагивает в первую очередь эпителиальный и эндотелиальный компартменты и запускает активацию каскада свертывания крови с образованием сгустка крови и высвобождение провоспалительных медиаторов, инициирующих инфильтрацию лейкоцитов.В качестве клеточных компонентов воспаления при заживлении кожных ран лейкоциты являются не только эффекторными клетками, регулирующими иммунные ответы, но также участвуют в анаболической фазе деградации тканей за счет выработки протеаз и реактивных промежуточных соединений кислорода, и особенно в катаболической фазе ткани. ремоделирование за счет производства факторов роста. Рекрутинг лейкоцитов пространственно-временной и дифференцированно регулируется хемокинами. Более того, присутствие хемокиновых рецепторов на резидентных клетках, таких как кератиноциты и эндотелиальные клетки, указывает на то, что хемокины также способствуют регуляции эпителизации, ремоделирования тканей и ангиогенеза (3).Таким образом, хемокины занимают исключительную позицию в плане интеграции воспалительных явлений и репаративных процессов (4). В дополнение к хемокинам, процесс восстановления выполняется и стимулируется не менее сложной сигнальной сетью, которая включает в себя многочисленные факторы роста и цитокины, которые изменяют рост, дифференцировку и метаболизм клетки-мишени с помощью паракринных, аутокринных, юкстакринных или эндокринных механизмов. Эффекторные клетки ведут себя как следствие их связывания со специфическими поверхностными рецепторами или белками внеклеточного матрикса, чтобы запустить каскад молекулярных событий, который приводит к регуляции специфических генов для контроля клеточного цикла, подвижности или паттернов дифференцировки (5).Использование трансгенных мышей и мышей с нокаутом дает интересные и часто неожиданные результаты, которые показывают in vivo функцию медиаторов при заживлении ран (6). Однако, несмотря на описание роли TCRγ (7), TNFRp55 (8), хемокинового рецептора CXCR2 (9) и CX3CR1 (10), все еще имеется мало информации о родственных рецепторах, особенно о TLR, отвечающих на эти медиаторы в заживление кожных ран.
TLR относятся к семейству рецепторов распознавания образов, которые распознают различные молекулярные паттерны, связанные с патогенами, включая молекулы грамположительных и -отрицательных бактерий, ДНК- и РНК-вирусов, грибов, простейших и различных агонистов, происходящих от хозяина (11 ).Эти рецепторы составляют первую линию защиты от патогенов и играют решающую роль во врожденной иммунной системе за счет выработки воспалительных цитокинов и хемокинов (12). Передача сигналов TLR также участвует в развитии адаптивных иммунных ответов за счет активации костимулирующих молекул на APC (11). Предыдущее исследование продемонстрировало заметно более медленное заживление эксцизионных кожных ран у мышей с дефицитом MyD88 (13). MyD88 является сигнальным адаптером для всех TLR, кроме TLR3, который передает сигнал через альтернативную молекулу адаптера, содержащий домен Toll / IL-1R, адаптер, индуцирующий IFN-β (TRIF) (14).Экспрессия TLR3 была продемонстрирована ранее в здоровой и псориатической коже (15), а также в культивированных эпидермальных кератиноцитах нормальной кожи (16). Новые данные показывают, что РНК, высвобождаемые либо из поврежденной ткани, либо внутри эндоцитированных клеток, служат эндогенными лигандами для TLR3, чтобы индуцировать TNF-α и усиливать экспрессию TLR3 в кератиноцитах (17, 18). У мышей с дефицитом TLR3 воспаление уменьшилось с более низкой экспрессией IL-6 и TNF-α на краю раны через 3 дня асептического повреждения (18). Таким образом, TLR, по-видимому, регулируют воспалительные реакции во время заживления кожных ран.
В этом исследовании мы исследовали роль TLR3 в модели заживления ран у мышей. Множественные параметры раны, включая инфильтрацию воспалительных клеток, экспрессию хемокинов и их рецептор, оценивали в различные интервалы времени. Мы обнаружили, что у мышей TLR3 — / — наблюдается замедленное заживление ран с нарушением неоваскуляризации и нарушением рекрутирования нейтрофилов и макрофагов, связанным со сниженной экспрессией хемокинов MIP-2 / CXCL2, MIP-1α / CCL3 и MCP-1 / CCL2 в ранах. .
Материалы и методы
Реагенты и Abs
Ab козьих антител против IgG-пероксидазы кролика, Ab козьих антител против IgG-пероксидазы крыс и Ab козьих антител против IgG-пероксидазы кроликов были приобретены у Sigma (Сент-Луис, Миссури) . Реагент TRIzol и Hoechst 33342 были от Invitrogen (Карлсбад, Калифорния). Набор реагентов ExScript RT был от TaKaRa (TaKaRa Biotechnology, ДаЛиан, Китай). Смесь Brilliant SYBR Green QPCR Master Mix была произведена в Stratagene (La Jolla, CA). FITC-конъюгированные антимышиные F4 / 80 и FITC-конъюгированные антимышиные CD3 были от eBioscience (Сан-Диего, Калифорния).Поликлональные антитела кролика против YM1 мыши были получены от StemCell Technologies (Ванкувер, Британская Колумбия, Канада). Кроличьи антимышиные FIZZ1 были от PeproTech (Лондон, Великобритания). Кроличий антимышиный кератин / цитокератин Ab был от Nichirei Biosciences (Chuo-ku, Tokyo, Japan). Cy3-конъюгированные ослиные антимышиные и поликлональные антитела против козьего IgG были приобретены в Jackson ImmunoResearch Laboratories (West Grove, PA). Кроличьи антимышиные TLR3, козьи антимышиные PECAM-1 (CD31), козьи антимышиные миелопероксидазы (MPO), мышиный антимышиный α-актин гладких мышц ([SMA], который перекрестно реагирует с мышиным α-SMA), Конъюгированные с FITC ослиные анти-козьи, козьи поликлональные антитела против кроличьих IgG, контрольные IgG изотипа или преиммунные сыворотки были приобретены в Santa Cruz Biotechnology (Санта-Крус, Калифорния).Субстрат-хромоген диаминобензидина был от Dako Cytomation (Carpinteria, CA). Наборы ELISA для мышиных MIP-2 / CXCL2, MIP-1α / CCL3 и MCP-1 / CCL2 были от R&D Systems (Миннеаполис, Миннесота). Набор для быстрого старта Брэдфордского протеинового анализа ELISA был от Bio-Rad Laboratories (Геркулес, Калифорния). Большинство других реагентов, таких как компоненты соли и буфера, были аналитической чистоты и были получены от Sigma.
Животные
TLR3 — / — мышей были приобретены у Taconic Farms. Мышей подвергали обратному скрещиванию ≥10 поколений на фон C57BL / 6, а затем скрещивали для получения однопометников с нокаутным генотипом и дикого типа (WT) (в качестве контроля).Во всех экспериментах использовали однопометников обоего пола в возрасте от 8 до 12 недель. Животных содержали индивидуально в клетках в определенных условиях, свободных от патогенов, и им давали воду и стандартную лабораторную пищу ad libitum во время экспериментов. Генотипирование животных проводили с помощью ПЦР ДНК, полученной из биоптатов хвоста. Праймеры были синтезированы в соответствии с протоколом генотипирования (инвентарный номер 005217) из лаборатории Джексона (Бар-Харбор, штат Мэн). Уход за животными и все экспериментальные процедуры были одобрены институциональным комитетом по уходу и использованию лабораторных животных.
Модель раны in vivo
Полнослойные раны были созданы на дорсальной коже в стерильных условиях. Вкратце, мышей анестезировали i.p. прием Кессодрата (хлоралгидратов). После депиляции 10% Na 2 S и очистки повидон-йодом (Бетадин) и 70% этанолом, дорсальную кожу собирали по средней линии и пробивали через два слоя кожи стерильным одноразовым биопсийным перфоратором (4 мм в диаметре). диаметр; Miltex), образуя по одной ране с каждой стороны средней линии.Та же процедура была повторена, получив четыре раны. Каждый участок раны был сфотографирован в цифровом виде через указанные интервалы времени, а площади раны были определены на фотографиях с использованием Adobe Photoshop (версия 7.0; Adobe Systems) (рис. 2). Изменения площади раны с течением времени выражали в процентах от первоначальной площади раны.
Гистопатологический и иммуногистохимический анализы
Образцы ран фиксировали в 4% формальдегиде и затем заливали парафином. Срезы окрашивали H&E для гистологического анализа.Иммуногистохимические анализы проводили с использованием анти-TLR3. Срезы инкубировали с Ат в концентрации 0,5–5 мкг / мл при 4 ° C в течение ночи. После инкубации с вторичным Ab, конъюгированным с пероксидазой, добавляли тетрагидрохлорид диаминобензидина хромогена. После того, как цвет достаточно развился, срезы контрастировали гематоксилином Майера.
Иммунофлуоресцентный анализ
Двухцветный иммунофлуоресцентный анализ выполняли для определения типов клеток, экспрессирующих TLR3, в раненой коже.Срезы инкубировали с комбинацией анти-TLR3 и -кератина, -F4 / 80, -α-SMA, -CD31, -MPO или IgG изотипического контроля при 4 ° C в течение ночи. После инкубации с соответствующими вторичными антителами, конъюгированными с флуорохромом, и контрастирования с помощью Hoechst 33342, срезы были закреплены и визуализированы на флуоресцентном микроскопе Zeiss (визуализация Axioplan 2; Carl Zeiss MicroImaging GmbH, Германия) с многоканальным режимом и фильтрами Zeiss (набор 17 для FITC: полоса пропускания 515–565; установка 20 для Cy3: полоса пропускания 575–640; и установка 02 для Hoechst 33342: LP 420).Срезы также окрашивали антителами к CD31 для оценки неоваскуляризации.
Лейкоцитарная инфильтрация
Ложе раны было определено как область, окруженная неповрежденной кожей и фасциями, регенерированным эпидермисом и струпом. Замороженные криостатные срезы тканей кожной раны иммуноокрашивали антителами против МПО, -F4 / 80 и -CD3 для мечения нейтрофильных гранулоцитов, макрофагов и Т-клеток соответственно; другие срезы дополнительно обрабатывали для двухцветного иммунофлуоресцентного анализа для идентификации типов макрофагов YM1 или FIZZ1 + , затем срезы контрастировали с помощью Hoechst 33342.Для статистического анализа количество типов инфильтрирующих клеток в ложе раны было подсчитано с помощью программного обеспечения Image-Pro Plus в 10 случайно выбранных полях зрения (увеличение × 400) срезов и среднее количество (± SD) клеток в Было рассчитано 10 полей.
ELISA
Образцы ран вырезали и гомогенизировали в 0,4 мл лизирующего буфера (10 мМ PBS, 0,1% SDS, 1% Nonidet P-40 и 5 мМ EDTA), содержащего ингибиторы протеаз. Гомогенаты центрифугировали при 12000 об / мин в течение 15 мин.Супернатанты использовали для определения уровней MIP-2 / CXCL2, MIP-1α / CCL3 и MCP-1 / CCL2 с помощью коммерческих наборов для ELISA в соответствии с инструкциями производителя. Общий белок в супернатантах измеряли по методу Брэдфорда. Данные были выражены как количество целевой молекулы (пг) на общий белок (мг) для каждого образца.
Выделение тотальной РНК и количественная ПЦР в реальном времени
Общую РНК выделяли из неповрежденных и раненых образцов кожи с использованием набора реагентов TRIzol, а кДНК получали обратной транскрипцией.Экспрессию мРНК тестируемых молекул в образцах кожи определяли с помощью ПЦР в реальном времени в соответствии с протоколом производителя. Каждая реакция содержала 12,5 мкл 2 × SYBR Green Master Mix, 300 нМ олигонуклеотидных праймеров (таблица I), синтезированных Invitrogen Biotechnology, (Шанхай, Китай), 10 мкл кДНК в разведении 1:10 и воды до общего объема 25. мкл. Условия термоциклирования включали начальную денатурацию при 95 ° C в течение 10 минут, 40 циклов при 95 ° C в течение 30 с, 60 ° C в течение 1 минуты и 72 ° C в течение 1 минуты.Концентрации закодированных образцов рассчитывались с использованием программного обеспечения (версия 1.7), поставляемого с системой ABI 7700 (Perkin Elmer Applied Systems, Foster City, CA).
Синтез и обработка малой интерферирующей РНК
Последовательности двух олигонуклеотидных цепей дуплекса малой интерферирующей РНК (миРНК) TRIF следующие: смысл 5′-GCUAUGUAACACACCGCUGTT-3 ‘; и антисмысловой 5’-CAGCGGUGUGUUACAUAGCTT-3 ‘. Эта целевая последовательность siRNA присутствует в мРНК TRIF для мыши (Справочная последовательность Национального центра биотехнологической информации: NM_174989.3) ген. TRIF-специфическая миРНК и скремблированная контрольная миРНК были синтезированы Ribobio (Гуанчжоу, Китай). Комплементарные смысловые и антисмысловые одиночные цепи отжигали, обессоливали и лиофилизировали, чтобы получить конечный дуплекс миРНК. Целостность и чистота миРНК были подтверждены методами ВЭЖХ и масс-спектроскопии MALDI-TOF. Лиофилизированный дуплекс суспендировали в буфере PBS без РНКазы-ДНКазы перед использованием in vivo. После операции на ране, 5 нмоль специфической TRIF-siRNA в 20 мкл стерильного раствора PBS без эндотоксина наносили локально на две раны на правой стороне мышей ежедневно, тогда как другие две раны на левой стороне дорсальной кожи получали такой же объем контроля. скремблированная миРНК или PBS соответственно.Каждая группа состояла из восьми мышей. Макроскопическую площадь открытой раны измеряли и анализировали через указанные интервалы времени. Для оценки эффективности сайленсинга TRIF раненые участки кожи, обработанные миРНК в течение 5 дней, были разрезаны, общая РНК была выделена, и количественная ПЦР в реальном времени была проведена с TRIF-специфическими праймерами, как показано в таблице I. Уровни экспрессии мРНК в каждом Образец определяли после нормализации относительно уровня мРНК GAPDH.
Воспроизводимость и статистический анализ
Все эксперименты повторяли как минимум дважды.На всех рисунках показаны объединенные данные повторных экспериментов или репрезентативных экспериментов, как указано. Данные выражены как среднее значение ± стандартная ошибка среднего для указанного количества независимо выполненных дублированных экспериментов. Статистическая значимость между средними значениями была проанализирована с помощью двустороннего критерия Стьюдента t с использованием версии SPSS 13.0. Статистически значимым считалось значение p <0,05.
Результаты
Отсроченное заживление ран у мышей TLR3
— / —Было показано, что TLR3 необходим для нормальных воспалительных реакций после травмы (18).Исследования также показали, что активация TLR3 ингибирует ангиогенез (19, 20). Эти результаты предполагают, что TLR3 играет важную роль в заживлении ран. Чтобы проверить участие TLR3 в повреждении кожи, мы сначала оценили изменения в ране у мышей с дефицитом TLR3. Четыре круглые иссеченные раны на всю толщину кожи были созданы симметрично на спине мышей (рис. 1 A ). Области ран анализировали на протяжении всего процесса заживления (рис. 2). У однопометников из контрольной группы раны постепенно и прогрессивно заживали, достигая полного заживления к 12-м суткам после травмы (рис.1 В ). К нашему удивлению, закрытие раны было заметно задержано, а не ускорено у мышей TLR3 — / — (Fig. 1 B ). На 7 день после ранения контрольные раны уже утратили свои струпья и выглядели полностью эпителизированными, тогда как раны мышей TLR3 — / — показали только частичную эпителизацию и все еще несли струп (фиг. 1 B ). Статистический анализ показал, что закрытие раны у мышей TLR3 — / — было значительно задержано по сравнению с контрольными мышами, начиная с 5-го дня после травмы (рис.1 С ). Полное закрытие раны было достигнуто на 14 день после операции у мышей TLR3 — / — с задержкой на 2 дня по сравнению с контрольными однопометниками (фиг. 1 B , 1 C ). Гистологическая оценка показала, что раны у мышей TLR3 — / — имели более тонкие и короткие неоэпидермальные пласты на краю раны (фиг. 1 D ). Кроме того, реэпителизация, образование грануляционной ткани и неоваскуляризация были уменьшены, а эпителиальный промежуток увеличился в ранах у мышей TLR3 — / — по сравнению с контрольными однопометниками на 5-й день после травмы (рис.1 D ). Наши данные продемонстрировали, что у мышей с дефицитом TLR3 заживление ран замедлялось.
РИСУНОК 1.Отсроченное заживление ран у мышей TLR3 — / — . A , Полнослойные кожные раны были созданы у мышей WT или TLR3 — / — . B , Репрезентативные фотографии мышей WT и TLR3 — / — , показывающие макроскопическое закрытие раны в разные дни после травмы. C , Микрофотографии ран анализировали через указанные интервалы времени для определения закрытия у мышей WT и TLR3 — / — .Все значения представляют собой среднее значение ± SEM. * p <0,05 указывает на достоверность, а ** p <0,01 указывает на очень значительно более высокие значения, наблюдаемые у мышей WT по сравнению с мышами TLR3 — / — . D , гистопатология ( левая и средняя панели ) и иммуногистохимия (окрашивание CD31, правые панели ) ран мышей WT и TLR3 — / — на 5 день после травмы. Мыши TLR3 — / — показали меньшую реэпителизацию, образование грануляций ( нижняя левая и средняя панели ) и неоваскуляризация ( нижняя правая панель ). Рамки на левых панелях увеличены на средних панелях . Многослойный неоэпидермис был виден на краю ран у мышей WT, тогда как эпителиальная ткань была дезорганизована в ранах TLR3 — / — . Показаны репрезентативные результаты трех независимых экспериментов с четырьмя животными в каждой группе. Исходное увеличение × 40 ( левые панели ) и × 400 ( средние и правые панели ).
РИСУНОК 2.Измерение и анализ ран.В день 0 на коже спины мышей WT или TLR3 — / — были созданы четыре круглые эксцизионные раны (диаметром 4 мм) на коже спины. Каждый участок раны был сфотографирован в цифровом виде через указанные интервалы времени, а площади раны были определены на фотографиях с использованием программного обеспечения Adobe Photoshop. Изменения площади раны с течением времени выражали в процентах от первоначальной площади раны.
Снижение инфильтрации лейкоцитов в кожные раны у мышей TLR3
— / —Затем мы исследовали набор лейкоцитов, критическую процедуру заживления ран, в местах ран.Нейтрофилы, макрофаги и Т-лимфоциты были окрашены иммунофлуоресценцией анти-МПО (фиг.3 A ), анти-F4 / 80 (фиг.3 B ) и анти-CD3 Abs (фиг.3 C ). ), соответственно. Накопления нейтрофилов и макрофагов в нормальной коже не было (данные не показаны). Приток нейтрофилов достиг пика в 1 день и постепенно снижался, по крайней мере, до 6 дня во всех ранах (фиг. 3 D ). Макрофаги F4 / 80 + начали накапливаться на 1-й день после травмы и достигли максимального уровня на 6-й день (рис.3 E ). Кинетика инфильтрации лейкоцитов в раны была сходной у контрольных TLR3 и — / — животных (фиг. 3 D , 3 E ). Однако инфильтрация нейтрофилов в раны мышей TLR3 — / — была значительно снижена в дни 1 и 3 (фиг. 3 D ). Рекрутирование макрофагов в раны у мышей TLR3 — / — было значительно ослаблено на 3 и 6 дни по сравнению с контрольными однопометниками (фиг. 3 E ). Уровень инфильтрации Т-лимфоцитов CD3 + был сходным у однопометников TLR3 — / — и WT (рис.3 Ф ). Затем мы исследовали инфильтрацию альтернативно активированных макрофагов / M2, субпопуляции макрофагов, которые вносят вклад в заживление ран (19, 21-25). Маркеры этих макрофагов, FIZZ1 / RELMα и YM1, использовали для окрашивания срезов ран. Мы обнаружили, что через 6 дней после травмы в ткани раны как в контрольной, так и в TLR3 — / — были многочисленные двойные положительные макрофаги F4 / 80 и YM1 или FIZZ1. Однако количество ранозаживляющих макрофагов YM1 + или FIZZ1 + в ранах мышей TLR3 — / — было заметно снижено по сравнению с ранами мышей дикого типа (рис.4).
РИСУНОК 3.Снижение набора лейкоцитов в ране мышей TLR3 — / — . A – C , Иммунофлуоресцентный анализ клеточной инфильтрации с анти-МПО ( A ), анти-F4 / 80 ( B ) или анти-CD3-антителами ( C ) или контрольными IgG-антителами в кожной ране срезы мышей WT и TLR3 — / — . Оригинальное увеличение × 400. D – F , Накопление нейтрофилов ( D ), макрофагов ( E ) и Т-лимфоцитов ( F ) в ранах мышей WT и TLR3 — / — .Все значения представляют собой среднее значение ± SEM. * p <0,05, значительно более высокие значения наблюдались в ранах мышей WT. Показаны репрезентативные результаты трех независимых экспериментов с четырьмя животными в каждой группе.
РИСУНОК 4.Сниженное привлечение макрофагов YM1 + ( A ) и FIZZ1 + ( B ) в рану мышей TLR3 — / — . Ткани раны на 5 день мышей — / — WT и TLR3 дважды окрашивали на F4 / 80 (зеленый) и YM1 или FIZZ1 (красный).Сигналы были объединены в цифровом виде в правых панелях. Для отрицательных контролей ткани окрашивали контрольными антителами IgG и контрастировали с помощью Hoechst 33342. Исходное увеличение × 400. Показаны репрезентативные результаты трех независимых экспериментов с четырьмя животными в каждой группе.
Снижение экспрессии хемокинов в кожных ранах мышей TLR3
— / —Заметное снижение инфильтрации нейтрофилов и макрофагов в ранах мышей TLR3 — / — предполагало возможность экспрессии хемокинов.Чтобы проверить эту гипотезу, мы использовали количественные ПЦР для изучения изменений в экспрессии мРНК хемокинов (таблица I). Все 15 протестированных генов хемокинов были активированы в ранах в различной степени с различной кинетикой (данные не показаны), что согласуется с предыдущими сообщениями (4, 6, 8, 10, 26–29). Однако экспрессия генов MIP-2 / CXCL2, MIP-1α / CCL3 и MCP-1 / CCL2 была значительно снижена у мышей TLR3 — / — по сравнению с контролем WT (фиг. 5 A ). MIP-2 / CXCL2 снизился на 7.Пятикратно в первый день повреждения кожи у мышей TLR3 — / — . После этого уровень мРНК MIP-1α / CCL3 и MCP-1 / CCL2 преимущественно снижался у мышей TLR3 — / — . Соответственно, уровень белка этих хемокинов был значительно повышен в ранах мышей WT (фиг. 5 B – D ). Напротив, уровень белка MIP-2 / CXCL2, MIP-1α / CCL3 и MCP-1 / CCL2 в кожных ранах TLR3 — / — был значительно снижен по сравнению с мышами WT (рис. 5 B – D ). ).Эти результаты предполагают, что активация TLR3 может индуцировать продукцию хемокинов, которые необходимы для набора лейкоцитов в раны.
Таблица I. Последовательности праймеров для ПЦР в реальном времени. Рисунок 5.Экспрессия хемокинов в кожных ранах. A , ПЦР-анализ в реальном времени экспрессии мРНК хемокинов в ранах на 1, 3 и 5 дни после травмы у мышей WT и TLR3 — / — . Относительные изменения были нормализованы относительно мРНК GAPDH и рассчитаны с использованием метода 2 -ΔΔ CT.Результаты были из трех отдельных экспериментов, выполненных в двух экземплярах, и выражены как среднее ± SEM кратного увеличения по сравнению с контролем. * p <0,05 указывает на значительно более высокие значения у мышей WT по сравнению с исходным уровнем у мышей TLR3 — / — . B – D , Уровни белка MIP-2 / CXCL2 ( B ), MIP-1α / CCL3 ( C ) и MCP-1 / CCL2 ( D ) в ранах WT и TLR3 — / — мышей измерено с помощью ELISA. Значения представляют собой среднее значение ± стандартная ошибка среднего повторных определений.* p <0,05 указывает на значительно более высокие значения у мышей WT. Показаны репрезентативные результаты трех независимых экспериментов с четырьмя животными в каждой группе.
Экспрессия TLR3 при заживлении ран
Количественная ПЦР показала, что экспрессия мРНК TLR3 была значительно повышена в раненой коже мышей WT, наиболее сильно в день 1 (фиг. 6 A ). Иммуногистохимический анализ показал, что, хотя белок TLR3 обнаруживался как в дерме, так и в подкожной клетчатке нормальной кожи, он значительно усиливался после повреждения кожи (рис.6 В ). Большое количество клеток TLR3 + было обнаружено в ране и прилегающих областях, которые оставались повышенными до 6 дней после ранения (данные не показаны). Эксперименты по совместной локализации показали, что клетки TLR3 + были резидентными клетками (кератин + кератиноцитов, α-SMA + фибробласты и CD31 + эндотелиальные клетки) и инфильтрирующими лейкоцитами (MPO + + нейтрофилов + F4 макрофагов) в ранах кожи (рис.6 С ). Эти результаты предполагают, что активация TLR3 эндогенными лигандами из поврежденной ткани может регулировать нормальное заживление ран.
РИСУНОК 6. ЭкспрессияTLR3 в коже. A , ПЦР-анализ экспрессии TLR3 в кожных ранах в реальном времени. Анализировали соотношение TLR3 и GAPDH на участках ран через 1, 3 или 5 дней после ранения. Результаты были получены из трех отдельных экспериментов, которые были выполнены в двух экземплярах и выражены как среднее ± SEM кратного увеличения по сравнению с контролем. * p <0.05 указывает на значительно более высокие значения по сравнению с нормальной кожей. B , Иммуногистохимический анализ экспрессии TLR3 в дерме (, левая панель, ) и подкожной ткани (, правая панель, ) нормальной кожи ( верхние панели, ) или раненой коже ( нижние панели, ) на 1 день после травмы. C , Клеточный источник экспрессии TLR3 на участке раны. Двухцветные иммунофлуоресцентные изображения показаны на 3-й день после травмы с использованием антицитокератина, анти-CD31, анти-α-SMA, анти-F4 / 80 или анти-MPO (зеленый) и анти-TLR3 (красный) Abs.Сигналы были объединены в цифровом виде в правых панелях. Для контроля срезы окрашивали контрольными антителами или антителами TLR3 и контрастировали с помощью Hoechst 33342. Показаны репрезентативные результаты трех независимых экспериментов. Оригинальное увеличение × 400.
Влияние подавления миРНК TRIF на заживление ран
TRIF является критическим адаптерным белком для TLR3-опосредованной передачи сигналов (30). Чтобы прояснить роль передачи сигналов TLR3 в заживлении кожных ран, мы исследовали эффект подавления TRIF.TRIF-специфическую или контрольную миРНК наносили местно на раны. Применение TRIF-специфической миРНК значительно задерживало закрытие раны у мышей WT по сравнению с лечением PBS, тогда как введение контрольной скремблированной миРНК значительно улучшало заживление ран у мышей WT (фиг. 7 A , 7 B ). Молчание TRIF с помощью TRIF-специфической миРНК было подтверждено анализом ПЦР в реальном времени, который показал, что экспрессия мРНК TRIF на 5 день была значительно снижена по сравнению с ранами, обработанными скремблированной миРНК или PBS (рис.7 С ). Мы также исследовали влияние скремблированной миРНК на TLR3 — / — ран. Данные показали, что обработка скремблированной миРНК не улучшила заживление ран у мышей TLR3 — / — в течение 14 дней (рис. 7 D , 7 E ), что свидетельствует о роли TLR3 в опосредовании лучшего эффекта. скремблированной siRNA, чем PBS, при заживлении ран (фиг. 7 B ). Взятые вместе, эти результаты дополнительно демонстрируют важную роль оси TLR3-TRIF в процессе заживления ран после повреждения кожи.
РИСУНОК 7.Отключение TRIF замедляет заживление кожных ран у мышей WT. Пять наномолей TRIF-siRNA в 20 мкл стерильного PBS наносили местно на две раны, тогда как другие раны получали такой же объем PBS или контрольную скремблированную siRNA. A , Макроскопический вид заживления раны, обработанной исследуемыми растворами, в разные дни после травмы. Показаны репрезентативные результаты трех независимых экспериментов с шестью животными. B , В указанные моменты времени площадь раны определяли с использованием анализа изображений и выражали в процентах от исходной площади раны.Значения представляют собой среднее значение ± стандартная ошибка среднего. * p <0,05 по сравнению с контрольной обработкой скремблированной миРНК. † p <0,05 по сравнению с лечением PBS. Показаны репрезентативные результаты трех независимых экспериментов. C , уровень мРНК TRIF в ранах, подавляющих TRIF. Тотальную РНК выделяли из ран, ежедневно обрабатываемых TRIF-специфической миРНК, скремблированной контрольной миРНК или носителем на 5-й день, и проводили количественную ПЦР в реальном времени. Результаты были получены из трех отдельных экспериментов, которые были выполнены в двух экземплярах и выражены как среднее ± SEM индукции складки по сравнению с исходным уровнем неповрежденной кожи. * p <0,05 по сравнению с контрольной обработкой скремблированной миРНК. † p <0,05 по сравнению с лечением PBS. D и E , Эффект скремблированной миРНК на заживление ран в TLR3 — / — ран. Раны мышей TLR3 — / — обрабатывали скремблированной миРНК и определяли площади ран. Показаны репрезентативные изображения ран ( D ) и кинетики закрытия раны ( E ) из трех независимых экспериментов.Значения представляют собой среднее значение ± стандартная ошибка среднего. * p <0,05 по сравнению с ранами, обработанными PBS.
Обсуждение
Наше исследование показало, что заживление кожных ран у мышей TLR3 — / — задерживалось в связи со сниженным накоплением нейтрофилов и альтернативно активированными макрофагами / M2 в сочетании со сниженной экспрессией хемокинов в ранах. Первоначально TLR3 был идентифицирован как специфический рецептор вирусной дцРНК (31). Однако недавние исследования показали, что TLR3 также распознает эндогенные РНК и играет важную роль в воспалительных реакциях на животных моделях перевязки слепой кишки, пункционной хирургии и разреза кожи (18, 32).Отчеты показали, что активация TLR3, запускаемая РНК, высвобождаемой некротическими клетками, индуцирует экспрессию TNF-α и TLR3 в кератиноцитах. У мышей TLR3 — / — наблюдалось снижение продукции провоспалительных цитокинов и рекрутирования лейкоцитов при экспериментальном колите (33) и остром повреждении кожи (18). Эти данные показывают, что TLR3 необходим для нормального воспалительного ответа после травмы. В соответствии с этими наблюдениями, наше исследование показало, что кожные раны усиливают экспрессию TLR3 у мышей WT.Врожденный ответ резидентных и рекрутированных клеток борется с вторжением микробов, но также может критически влиять на процесс восстановления, высвобождая широкий спектр медиаторов. Однако влияние воспаления на процесс заживления ран спорно (34). Анализ заживления ран на ряде моделей мышей с дефицитом индивидуальных медиаторов воспаления, рецепторов или истощения лейкоцитов дал разные результаты. Заживление ран ускоряется у мышей с дефицитом TNFRp55 (8), IFN-γ (35) или у мышей с истощением нейтрофилов (36), но нарушается у мышей с дефицитом TNF-α (37), MCP-1 (38). ) или Ил-6 (39).Наши результаты показывают, что врожденная воспалительная реакция улучшает процесс восстановления в коже, что будет полезно для дальнейшего понимания сложного и, возможно, тканезависимого процесса заживления ран.
Важность передачи сигналов TLR3 в заживлении кожных ран была дополнительно подтверждена молчанием TRIF. TRIF представляет собой адаптерный белок, содержащий домен Toll / IL-1R. Его сверхэкспрессия запускает TLR3-опосредованные сигнальные пути, что приводит к индукции цитокинов и хемокинов (30, 40).В этом исследовании молчание миРНК TRIF задерживает заживление ран (рис. 7). Напротив, скремблированная миРНК способствовала закрытию ран у мышей WT (фиг. 7), но не у мышей TLR3 — / — , что указывает на роль TLR3 в опосредовании эффекта скремблированной миРНК. Эти результаты согласуются с сообщениями о том, что системная доставка полиинозин-полицитидиловой кислоты, синтетического аналога вирусной дцРНК, улучшенное заживление ран (29) и неспецифическая siRNA также может запускать врожденный иммунитет через TLR3 (19). Эти данные показывают, что TLR3 может быть потенциальной терапевтической мишенью для лечения ран.
Хемокины являются важными медиаторами рекрутирования лейкоцитов. Мы обнаружили, что экспрессия MIP-2 / CXCL2 была снижена в ранах TLR3 — / — . Мышиный MIP-2 / CXCL2 известен как гомолог человеческого IL-8 (CXCL8). Сообщалось, что IL-8 способствует пролиферации кератиноцитов (41), созреванию грануляционной ткани (42) и неоваскуляризации (4). Наши данные о том, что экспрессия MIP-2 / CXCL2 была увеличена в ранах кожи мышей, предполагают важную роль, которую играет MIP-2 / CXCL2 в заживлении ран.У мышей GM-CSF KO отсроченное закрытие ран связывали со снижением MIP-2 / CXCL2, что вызывало уменьшение инфильтрации нейтрофилов и ангиогенеза (43). Исследование с использованием мышей, дефицитных по киназам семейства src (Hck и Fgr), продемонстрировало, что MIP-2 / CXCL2 индуцировал нейтрофил-зависимый ангиогенез in vivo (44). Таким образом, MIP-2 / CXCL2 может проявлять свою регулирующую активность непосредственно в отношении реэпителизации и неоангиогенеза или за счет своих хемоаттрактантных свойств нейтрофилов при заживлении ран. MCP-1 / CCL2 и MIP-1α / CCL3 представляют собой хемокины CC, привлекающие моноциты / макрофаги.Предыдущие исследования показали, что MCP-1 / CCL2 и MIP-1α / CCL3 были на высоких уровнях в ткани после ранения (6). Динамика экспрессии MCP-1 / CCL2 коррелировала с инфильтрацией макрофагов (6). Животные с нокаутом MCP-1 / CCL2 характеризовались значительно замедленной реэпителизацией раны и ангиогенезом (38). Обработка мышей нейтрализующей антисывороткой к MIP-1α / CCL3 перед травмой также уменьшала количество макрофагов в месте раны (6). Соответственно, наши данные показали, что как рекрутирование макрофагов, так и экспрессия MCP-1 / CCL2 и MIP-1α / CCL3 были увеличены у мышей WT.Следовательно, снижение количества макрофагов в ранах мышей TLR3 — / — может быть связано с дефектной экспрессией MIP-1α / CCL3 и MCP-1 / CCL2.
Предыдущие исследования in vitro показали, что продукция хемокина MIP-1α, усиленного лигандом TLR3 полиинозиновой-полицитидиловой кислотой, зависит от IFN типа I в кератиноцитах (28). Совсем недавно Грегорио и др. (45) показали, что инфильтрирующие плазматические дендритные клетки способствуют заживлению ран за счет продукции IFN типа I in vivo.В соответствии с этими наблюдениями мы также обнаружили, что экспрессия IFN-α и IFN-β в кожных ранах была повышена в кожных ранах мышей WT и TLR3 — / — , но в меньшей степени у TLR3 — / — мышей (данные не показаны). Необходимы дальнейшие исследования, чтобы выяснить, управляется ли продукция хемокинов in vivo за счет IFN-α / β после активации TLR3.
Фагоциты играют важную роль в заживлении ран (46). Нейтрофилы в ранах образуют первую линию защиты от местных инфекций и являются источником провоспалительных цитокинов, активирующих фибробласты и кератиноциты (47).Было высказано предположение, что нейтрофилы также могут участвовать в инициации ангиогенеза (46). Снижение количества нейтрофилов у мышей с дефицитом CXCR2 связано с нарушением заживления кожи (9). В соответствии с этими наблюдениями наши данные показали, что инфильтрация нейтрофилов в раны у мышей TLR3 — / — была уменьшена, что могло быть результатом снижения экспрессии MIP-2 / CXCL2. Макрофаги необходимы для нормального заживления ран. Они функционируют как фагоциты и APC и, в частности, являются важным источником факторов, критических для закрытия раны, таких как фактор роста эндотелия сосудов, фактор роста тромбоцитов, TGF-β, основной фактор роста фибробластов, гепарин-связывающий фактор роста эпидермиса, и TGF-α (1, 48, 49).Эти факторы стимулируют синтез внеклеточного матрикса локальными фибробластами, генерируют новые кровеносные сосуды, способствуют образованию грануляционной ткани и усиливают реэпителизацию (1, 48). Макрофаги также связываются с внеклеточным матриксом через свои интегриновые рецепторы для усиления фагоцитоза микроорганизмов и фрагментов внеклеточного матрикса, чтобы контролировать чистое содержание коллагена в ранах (48). Синтез нового коллагена, превышающий скорость разложения, приведет к фиброзу (50).Наше исследование показало уменьшение количества дважды позитивных альтернативно активированных / M2 макрофагов FIZZ1 и YM1 в ранах с отсроченным закрытием у мышей TLR3 — / — (51, 52), предполагая, что рекрутирование и дифференциация подмножества макрофагов может быть ключевым фактором. шаг в TLR3-опосредованном заживлении ран.
В совокупности, в этом исследовании мы показываем, что TLR3 важен для нормального заживления ран, на основании наблюдений на мышах TLR3 — / — и ингибирования передачи сигналов TLR3 у мышей WT с помощью siRNA, нацеленного на TRIF.Наши результаты показывают, что сигнальный каскад TLR3 может быть потенциальной терапевтической мишенью для аномального заживления ран при таких заболеваниях человека, как диабетическая стопа и цирроз печени.
Раскрытие информации
У авторов нет финансового конфликта интересов.
Благодарности
Мы благодарим доктора Джи Мин Ванга (Национальный институт рака, Национальные институты здравоохранения, Бетесда, Мэриленд) и доктора Ся Чжана (Колледж компьютерных, математических и естественных наук, Университет Мэриленда, Колледж-Парк, Мэриленд) за полезную критику рукописи.
Footnotes
Эта работа была поддержана Национальной программой фундаментальных исследований Китая (грант 2007CB512206) и Национальным фондом естественных наук Китая (грант 81072483).