Нелинейные деформаторы в Autodesk Maya
Деформаторы — это особые типы объектов, используемые для деформации формы других объектов, включая рассмотренные нами ранее полигональные и NURBS-объекты и широко применяемые как непосредственно в моделировании, так и в анимации. Плюсом их использования является возможность быстро изменять геометрию объекта без необходимости перемещения вручную его вершин.
Non Linear Bend Deformer Maya tutorial
За работу с деформаторами отвечает меню Deform (Деформировать), доступное только в режиме создания анимации (клавиша F2). Открывающийся через данное меню список деформаторов довольно велик — они находятся в подменю CreateNonlinear (Создать Нелинейные) и применяются для создания нелинейной деформации, отличающейся от линейной тем, что изменение формы объекта производится неравномерно вдоль его осей.
В Maya имеются следующие типы нелинейных деформаторов: Bend (Изгиб) — деформирует объект за счет его осевого изгиба;
- Flare (Выпуклость) — позволяет увеличить/уменьшить степень выпуклости объекта;
- Sine (Синусоидальная деформация) — деформирует объект за счет синусоидальных возмущений его поверхности в направлении одной из осей;
- Squash (Сплющивание) — обеспечивает сплющивание поверхности объекта в направлении одной из осей с сохранением его первоначального объема;
- Twist (Скручивание) — изменяет форму объекта путем применения деформации скручивания относительно одной из его осей;
- Wave (Волна) — деформирует объект за счет синусоидальных возмущений его поверхности в двух направлениях с образованием концентрических волн.

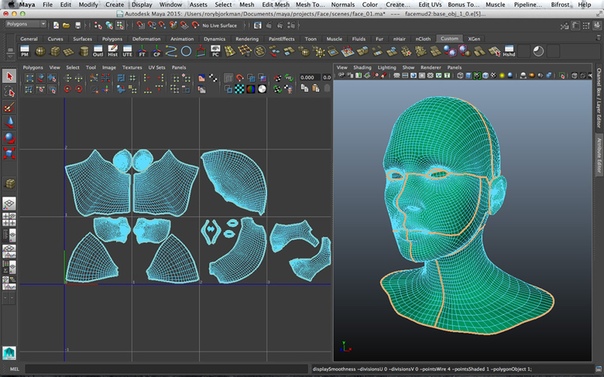
Назначенные объектам нелинейные деформаторы (как и прочие объекты) отображаются в окне Outliner (Структура), а результат применения любого из них определяется параметрами настройки деформатора (они регулируются обычным образом в окнах ChannelBox и AttributeEditor) и количеством сегментов самого объекта. Как правило, число сегментов устанавливается при создании объекта, однако при не совсем удачном применении деформатора оно может быть увеличено уже после его назначения, что, правда, может иметь негативные последствия — в таком случае приходится удалять деформатор, увеличивать степень разбиения объекта, а затем вновь его назначать. Как правило, при небольшом числе сегментов увеличение их количества приводит к изменению характера деформации, а при большом — способствует большей гладкости получаемой поверхности. При необходимости к объекту может быть применено сразу несколько деформаций — получаемый результат при этом чаще всего зависит от порядка следования деформаторов.
{fcomments}
3Dmaya.com.ua — Отзывы и рекомендации 3Dmaya
Содержание и ключевые слова
Важные и популярные веб-сайты
Веб-сайт с главной страницей «Главная Страница» также размещает информацию на страницах Лицензия Autodesk, Преподаватели и Работы Студентов. Ниже перечислены наиболее важныe подстраницы 3dmaya.com.ua :
| # | Описание | URL веб-сайта |
|---|---|---|
| 1. | Главная страница | / |
| 2. | Лицензия autodesk | /autodesk/ |
| 3. | преподаватели | /teachers/ |
| 4. | /students_works/ | |
5. | календарь занятий | /schedule/ |
| 6. | отзывы выпускников | /reviews/ |
| 7. | скачать видеоуроки | /maya_download/ |
| 8. | популярные вопросы | /questions/ |
| 9. | AR-creator | /sparkar/ |
| 10. | 3D Game Artist | /game-artist/ |
Информация о 3dmaya.com.ua из Wikipedia свободной энциклопедии: Autodesk Maya инструментальная система трёхмерной графики и компьютерной трехмерной .. узнать больше
Актуальные темы для 3dmaya.com.ua
отзывы о курсах 3dmaya.com.ua отзывы о 3dmaya.com отзывы курсов 3д майя харьков rhinoceros 3d
Ссылки на веб-сайт и присутствие в социальных сетях
В общей сложности найдено 4 указаний с других веб-сайтов на веб-сайт 3dmaya. com.ua. При этом наиболее распространенными понятиями в текстах ссылок являются курсы 3D и курсы. Наиболее важные ссылки на веб-сайт поступают от 3dpapa.ru и B3d.com.ua. Чаще всего на главную страницу ссылается 3dmaya.com.ua и на страницу
com.ua. При этом наиболее распространенными понятиями в текстах ссылок являются курсы 3D и курсы. Наиболее важные ссылки на веб-сайт поступают от 3dpapa.ru и B3d.com.ua. Чаще всего на главную страницу ссылается 3dmaya.com.ua и на страницу
Важнейшие ссылки на этот сайт
В таблице указаны наиболее важные ссылки на 3dmaya.com.ua, отсортированные по значимости в порядке убывания.
| # | Домен | Текст ссылки | Оценка ссылки |
|---|---|---|---|
| 1. | 3dpapa.ru | курсы 3D-моделирования, скульптинга, композитинга и анимации | |
| 2. | b3d.com.ua | курсы 3D графики |
Страницы, на которые наиболее часто ссылаются внешние источники
В таблице указаны веб-сайты 3dmaya. com.ua, на которые очень часто ссылаются другие веб-сайты, и которые, таким образом, составляют важную часть его содержания.
com.ua, на которые очень часто ссылаются другие веб-сайты, и которые, таким образом, составляют важную часть его содержания.
| # | URL веб-сайта | Количество внешних ссылок |
|---|---|---|
| 1. | http://3dmaya.com.ua/ (Главная страница) | 2 |
Замечание: Анализ внешних ссылок основан на самых последних полученных данных.
Профили и аккаунты в социальных сетях
В общей сложности в социальных сетях найдено два профилей для 3dmaya.com.ua, кроме того веб-сайт также представлен на Facebook и YouTube.
Техническая информация
Веб-сервер с IP-адресом 78.140.140.200 3dmaya.com.ua используется WebaZilla B.V. и расположен в Голландия. На данном веб-сервере обслуживается множество веб-сайтов. Оператор предоставляет данный веб-сервер в хостинг для многих клиентов. Преобладающим языком веб-сайтов является русский. Всего несколько веб-сайтов на данном веб-сервере не разрешены для детской аудитории.
На данном веб-сервере обслуживается множество веб-сайтов. Оператор предоставляет данный веб-сервер в хостинг для многих клиентов. Преобладающим языком веб-сайтов является русский. Всего несколько веб-сайтов на данном веб-сервере не разрешены для детской аудитории.Информация о сервере веб-сайта
| IP-адрес: | 78.140.140.200 |
| Оператор сервера: | WebaZilla B.V. |
| Количество сайтов: | более 100 — другие страницы с этим IP-адресом |
| Самые известные сайты: | Oslo.ru (известен), Dengi.fr (малоизвестен), 3dpapa.ru (малоизвестен) |
| Сайты для взрослых: | 2% веб-сайтов не разрешены для показа в детской аудитории |
| Языковое распределение: | 89% веб-сайтов русский, 4% веб-сайтов английский, 1% веб-сайтов итальянский |
Технические характеристики к технологии сайта
| ПО веб-сервера: | LiteSpeed |
| Cms-Программное обеспечение: | WordPress, Версия 5. 2.2 2.2 |
| Время загрузки: | 0,09 секунд (быстрее, чем 99 % всех сайтов) |
| HTML-версия: | HTML 5 |
| Размер файла: | 132,6 KB (1530 распознанных слов в сплошном тексте) |
Курс «3D моделирование в Autodesk Maya, профессиональный», Черкассы
Creativity is allowing yourself to make mistakes.
Design is knowing which ones to keep.
Scott Adams. 2013
В рамках данного курса Вы изучаете полный путь реализации объекта в 3D: полигональное и поверхностное моделирование, UV-сшивание, варианты текстурирования, освещение, задний план, подключение эффектов, финальный рендеринг. Для прохождения программы желательны базовые знания по графике.
Программное обеспечение:
- Autodesk Maya.
- Adobe Photoshop.
- библиотека HDRI Maps и Mental Ray текстур.

Краткая программа курса:
- Среда моделирования Autodesk Maya; определение, этапы, установка проекта.
- Интерфейс, настройка, установка панелей и пользовательских кнопок вызова функций.
- Навигация, переключение видов, панели атрибутов, манипуляторы, пивот.
- Работа с привязками, координатами, слоями, размерами, outliner.
- Полигональное проектирование. Функции Mash. Модификаторы полигонов Edit Mash.
- Архитектурные формы из полигонов. (Проект)
- Поверхностное проектирование. Функции Surfaces. Модификаторы поверхностей Edit Nurbs.
- Кривые. Способы построения и редактирование. Внедрение чертежей-профилей.
- Subdivisions, деформация, методы разбиения и детализации поверхностей.
- Скульптурная геометрия.
- Био-формы из поверхностей. (Проект)
- Динамика и nDynamics: Particles (воздух, дым, фракталы), Fluids (жидкости), Cloth, силовое поле, волосы
- Paint effect, Visor.
- Создание и редактирование временной анимации.
- Текстурирование: создание и настройка UV карты модели, подбор и сочетание шейдеров. Модуль hypershade. Шейдеры Mental Ray. Текстурирование развертки средствами Adobe Photoshop.
- Свет: выбор, расположение, настройка источников.
- Rendering: выбор, установка, настройки рендера в сочетании с шейдером и источником фотонов.
- Решение пространства, фона, переднего плана, глобального освещения; финальная сборка.
- Настройка эффектов каустики, окклюзии, отражения, свечения и пропускания.
- Камера, batch render.

Максимально детальное описание курса на сайте:
http://prestige-ua.mywebcommunity.org/03_3d.html
Viber, telegram, wechat, instagram, whatsApp
Facebook: #ourcoursesonline
полезные базовые функции для работы с 3Д моделями и их тестирования / Хабр
Недавно я столкнулся лицом к лицу с таким зверем как Autodesk Maya. За плечами у меня не было опыта в работе с 3Д моделированием и пришлось в экстренном порядке прокачиваться, начиная с основ, а также ознакомится с функциональностью данного ПО, чтобы проверить работу плагина (QA), который экспортил всё содержимое сцены в определенный формат. Эта статья рассчитана на людей без опыта либо с минимальным в Autodesk Maya и призвана познакомить вас с полезными функциями и редакторах, которые можно в ней найти. Если же вы не нашли функцию/редактор, который считаете полезным, то либо я с ним еще не работал, либо он не пригодился для моих нужд. Расскажите о них и о реальных случаях их применения в комментариях 🙂
Эта статья рассчитана на людей без опыта либо с минимальным в Autodesk Maya и призвана познакомить вас с полезными функциями и редакторах, которые можно в ней найти. Если же вы не нашли функцию/редактор, который считаете полезным, то либо я с ним еще не работал, либо он не пригодился для моих нужд. Расскажите о них и о реальных случаях их применения в комментариях 🙂
Статья разделена на секции, каждая из которых относится к одному из аспектов 3Д модели:
- Общие инструменты
- Работа с геометрией (полигональной сеткой)
- Работа с текстурами
- Работа с анимацией
- Shelf Editor, Shelves и немного о MEL (Maya Embedded Language)
1. Общие Инструменты
Данная секция содержит в себе общую информацию о Plug-in Manager, Outliner, Hypershade, Center Pivot, World Coordinate System и Working Units, а также Attribute Editor
1.1 Plug-in Manager
Если вам нужно установить плагин, то Plug-in Manager вам в помощь. Данный редактор содержит в себе список плагинов, находящихся в разных секциях, которые видит Maya. Каждая секция — это отдельный путь на файловой системе. Здесь нас интересуют 2 чекбокса: Loaded и Auto Load. Первый отвечает за загрузку плагина в данную сессию, второй же за загрузку плагина при открытии Maya.
Данный редактор содержит в себе список плагинов, находящихся в разных секциях, которые видит Maya. Каждая секция — это отдельный путь на файловой системе. Здесь нас интересуют 2 чекбокса: Loaded и Auto Load. Первый отвечает за загрузку плагина в данную сессию, второй же за загрузку плагина при открытии Maya.
Во избежания танцев с бубном из-за различных местоположений ваших плагинов, я советую класть ваши плагины в {MAYA_HOME}/bin/plug-ins
Открыть редактор можно из главного меню: Windows -> Settings/Preferences -> Plug-in Manager
1.2 Outliner
Outliner показывает иерархически список всех объектов в сцене. По умолчанию видны только DAG объекты (остальные скрыты по умолчанию) и не видны шейдеры, материалы и т.п… Для того, чтобы это изменить можно вызвать меню при помощи клика правой кнопкой мыши на Outliner и изменением статуса соответствующих чекбоксов. Пожалуй это самый используемый редактор в Maya, но ему в спину дышит редактор нод — Hypershade
1.
 3 Hypershade
3 HypershadeГлавный и основной редактор, если речь заходит о нодах. Hypershade является центральной рабочей областью для шейдинга. Здесь вы можете создавать редактировать и соединяя друг с другом различные ноды\узлы, такие как текстуры, материалы, источники света и т.д.
Открыть его можно:
- Нажав на иконку Hypershade в панели инструментов
- Из главного меню: Windows > Rendering Editors > Hypershade
Можно выделить следующие примеры использования:
- Удобно работать со всеми нодами в одном редакторе: легкое переключение между ними, добавление текстур, света, камер и т.п.
- Все свойства нод легко меняются в Property Editor.
- Вы можете сразу наблюдать эффект в при изменении материалов в Material Viewer абстрагируясь от вашей 3Д модели
1.4 World Coordinate System и Working Units
Вам может понадобится изменить вертикальную ось, а также меры измерений. Сделать это можно в Windows > Settings/Preferences > Preferences -> Settings
Сделать это можно в Windows > Settings/Preferences > Preferences -> Settings
Примеры использования:
- При импортие, чтобы убедиться, что значения и размеры объектов импортнулись в Maya корректно. Безусловно, метры в сантиметры и наоборот перевести не составит труда. Раскрывается данная функциональность если вы работаете, например с мало используемыми у нас футах, дюймах и т.п.
- При экспорте: если система, в которую вы, в последующем, будете импортить вашу сцену, должна принимать определенные измерительные единицы, например Unreal Engine 4.
1.5 Center Pivot
Функциональность, которая возвращает пивот (точку опоры) в центр элемента. Относительно этой точки пользователь делает все изменений (смещение, поворот, масштабирование). Крайне полезная функциональность, особенно если вы либо потеряли пивот, либо он находится в неположенном месте. Чтобы вернуть пивот в центр модели нужно использовать Modify -> Center Pivot. К слову, изменить местоположение пивота можно при помощи мыши при зажатой кнопки D на клавиатуре.
К слову, изменить местоположение пивота можно при помощи мыши при зажатой кнопки D на клавиатуре.
1.6 Attribute Editor
Attribute Editor хранит в себе все атрибуты выбранного объекта. Вкладки в верхней части позволяют выбирать узлы (nodes), подключенные к текущему. Здесь вы можете осуществлять все манипуляции с атрибутами ваших объектов. Незаменимая вещь для быстрого доступа к свойствам объекта, хотя большинство манипуляций можно совершать и из Hypershade. Открыть данные редактор можно при помощи Windows > General Editors > Attribute Editor / Windows > UI Elements > Attribute Editor, а также нажатием Ctrl+A (сочетание клавиш по умолчанию)
2. Работа с геометрией (полигональной сеткой)
Данная секция содержит в себе общую информацию о Poly Count, Normals, Tangents, Binormals, Triangulate/Quadrangulate mesh, Combine/Separate meshes, Select feature: object, face, vertex и другое, Blend objects (morphing), Reduce и Clean Up функциях.
Для последующей работы будет использована вкладка Modeling, проверьте, что у вас включен именно этот режим работы.
2.1 Poly Count
После включения в левом верхнем углу ViewPort у вас появится 3 столбца. В левом перечислены общие значения для всех видимых объектов, в среднем — значения для выбранных объектов, а в правом столбце показаны значения для выбранных компонентов объектов, которые частично видны во ViewPort. Чтобы включить Poly Count вам нужно выбрать «Display -> Heads Up Display —> Poly Count«
Примеры использования:
- Определение количества полигонов, треугольников, вертексов в модели просто по клику по 3д объекту или группе объектов
- Контроль количества полигонов во всей сцене
2.2 Включить отображение Normals, Tangents, Binormals
Функция включения отображения нормалей, тангентсов и бинормалей. Отображение включается на самой модели. Чтобы включить отображение вам нужно выделить объект и использовать Display -> Polygons -> Vertex Normals для нормалей и Tangents опцию для тангентсов и бинормалей в главном меню. Также можно использовать Face Normals опцию для отображения соответствующих нормалей.
Также можно использовать Face Normals опцию для отображения соответствующих нормалей.
Примеры использования:
- Крайне полезно для проверки этих атрибутов (в случае работы с группами сглаживания и т.п.)
2.3 Triangulate, Quadrangulate mesh
Функция триангуляции и квадрангуляции полигональной сетки меняет структуру полигональной сетки на треугольники (triangulate) или квадраты (quadrangulate). По умолчанию полигональная сетка в Maya состоит из квадратов. Данные функции находятся в меню Mesh: Mesh -> Triangulate/Quadrangulate
Примеры использования:
- Более точный прогноз финального количество вершин (vertices), граней и т.п.
- При триангулировании идет переиндексация, а та, в свою очередь, ускоряет обработку сетки.
2.4 Combine, separate meshes
Функция позволяющая объединять несколько полигональных объектов в один либо разделять на несколько. Использовать можно при помощи вызова из главного меню Mesh -> Combine/Separate
Примеры использования:
- Комбинирование объектов помогает сделать сложный объект из примитивов
- Разделите сложный объект на примитивы из которых он состоит, чтобы работать с отдельными основополагающими объектами.

2.5 Select: object, face, vertex и другое
Далеко не всегда нужно работать с целыми объектами. Если вам нужно выбрать отдельные элементы (грань, точку (vertex) и т.д.), то можете выбрать объект и кликнуть правой кнопкой мыши на Viewport и из контекстного меню выбрать что вы хотите выделять: Vertex, face, edge и т.д.
Примеры использования:
- Выделение отдельных частей сетки нужно для деформации объектов (вручную, функция Extrude и т.п.)и последующего морфинг (функция Blend Shape) и т.д.
- Удаление отдельных элементов в готовой сетке
2.6 Blend objects (morphing)
Смешивание объектов нужно для того, чтобы один объект трансформировался в другой. После применения данной функциональности можно контролировать степень трансформации, а также анимировать этот параметр. Чтобы активировать его вам нужно выбрать ваш основной объект и объект в который основной будет трансформироваться. Ваш основной объект должен быть выделен зеленым цветом. Чтобы активировать его вам нужно:
Чтобы активировать его вам нужно:
- Выделить целевой объект и его деформированные аналоги. Убедитесь, что объект, который будет подвержен трансформациям выделен зелёным цветом.
- Используйте Deform -> Blend Shape
- В Attribute Editor найдите вкладку Blend Shape и, используя соответствующие ползунки, регулируйте степень изменений.
Примеры использования:
- Трансформация одного объекта в другого полностью или частично
- Создание и использование анимации такой трансформации
2.7 Reduce (Poly Reduce)
Данная функциональность в автоматическом режиме уменьшает количество полигонов в вашей полигональной сетке. Чтобы использовать Reduce вам нужно выделить объект и использовать Mesh -> Reduce. Вы можете менять процент уменьшения полигонов, указать желаемое количество треугольников и т.п. после использования Reduce. Если у Maya не получится осуществить Reduce до желаемого вами значения, то она будет использовать ближайшее возможное от указанного вами.
Если ваша модель анимирована, то, после децимирования, анимация будет пересчитываться на каждом кадре при ее активации. Чтобы избежать этого вы можете удалить Non-Deformer History при помощи использования соответствующей функции (Edit -> Delete by Type -> Non-Deformer History), но тогда вы не сможете откатиться на предыдущие изменения при работе с вашей моделью, т.к. история будет удалена.
Стоит отметить, что данную функцию лучше не использовать на моделях со скелетной анимацией, т.к. в большинстве случаев, результат будет, мягко говоря, некорректен: деформации будут слишком сильные и ваша сетка будет расползаться из-за анимации.
Примеры использования:
- Уменьшение полигонов в сетке (как на всей, так и на ее части) для уменьшения размера файла, загрузки в нужные вам ресурсы и т.п.
2.8 Clean Up
Данная функциональность избавляется от ненужной вам геометрии в вашей полигональной сетке, такой как zero area faces, zero length edges, non-manifold mesh и т. п. Такая геометрия может появиться как при дизайне модели, так и после использования функции Reduce. Чтобы очистить модель вам нужно её выделить и выбрать Mesh -> Cleanup
п. Такая геометрия может появиться как при дизайне модели, так и после использования функции Reduce. Чтобы очистить модель вам нужно её выделить и выбрать Mesh -> Cleanup
Стоит отметить, что Cleanup сама может породить non-manifold геометрию, т.к. она объединяет вершины (vertices), убирает zero-length ребра и т.д. В таких случаях лучше используйте данную функциональность несколько раз.
Примеры использования:
- После использования Reduce бывают случаи, когда Maya отказывается экспортировать модели с анимацией и сама советует использовать Cleanup, чтобы убрать ненужную геометрию.
3. Работа с текстурами
Данная секция содержит в себе общую информацию о File Path Editor, UV Editor.
3.1 File Path Editor
Данный редактор хранит в себе все пути для ссылок, текстур, аудио и других файлов, которые включены в вашу сцену. File Path Editor позволяет быстро и легко диагностировать и исправлять проблемы с путями к файлам, например переназначить их.
Открыть редактор можно выбрав “Windows -> General Editors -> File Path Editor”
Примеры использования:
- Исправление путей, по которым уже не найти нужные файлы
- Замена одних файлов на другие
3.2 UV Editor
UV Editor позволяет просматривать и редактировать UV координаты текстур для полигонов, NURBS и т.д. Чтобы включить его вам нужно использовать “Windows -> Modeling Editors -> UV Editor” или через меню UV -> UV Editor в меню для моделирования (Modeling). В данном редакторе вы можете делать все необходимые манипуляции с UV координатами.
Примеры использования:
- Работа с UV координатами и текстурной картой, их модификация (flip modify и т.д.)
- Мгновенный просмотр итоговой текстуры и ее координат после применения всех модификаторов
- Просмотр ваших текстур в различных режимах: Wireframe, UV distortion и т.д.
4.
 Работа с анимацией
Работа с анимациейДанная секция содержит в себе общую информацию о Graph Editor, Time Editor, инструменте Quick Rig.
4.1 Graph Editor
Graph Editor хранит в себе список всех файлов сцены и, при клике на объекты, отображает их анимации, если таковые имеются. Внутри данного графика вы можете создавать, просматривать, изменять кривые анимации, ключевые кадры и т.п. Открыть Graph Editor вы можете из главного меню: “Windows -> Animation Editors -> Graph Editor”
Примеры использования:
- Создание, обновление, удаление ключевых кадров
- Изменение кривых анимаций и интерполяции\э между ними
- Работа с разными типами анимации: TRS (translation, rotation, scale), Skeleton, Morphing
4.2 Time Editor
Time Editor — это комплексный инструмент для создания и редактирования анимации. Он объединяет процессы редактирования анимации из существующих редакторов, предоставляя вам интуитивно понятный рабочий процесс на основе клипов для нелинейного редактирования анимации, который позволяет легко перемещаться между представлениями на уровне клипов и деталями на уровне клавиш.
Чтобы открыть редактор времени вам нужно главном меню Maya выбрать «Windows -> Animation Editors -> Time Editor.
Примеры использования:
- Создание и удобная работа с последовательностями анимаций у любого объекта или атрибута в Maya
- Группировка, обновление клипов
- Установка зоны воспроизведения анимаций
4.3 Инструмент Quick Rig
Quick Rig нужен, чтобы в автоматическом режиме связать вместе вашу модель и скелет.
Данный инструмент находится в панели “Animation” и крайне полезен, если вам не нужна высокая точность в весах, но нужна модель с привязанным к ней скелетом.
Чтобы открыть панель для работы с данным инструментом, вам нужно:
- Выбрать модель и скелет
- В панели “Animation” открыть “Windows -> Animation Editors -> Quick Rig”
- Нажать кноку “Auto-Rig”
Хотя, если вы в принципе не хотите возиться со скелетной анимацией, то можете скачать уже готовую модель с разными пресетами движениями на ресурсе Mixamo. Вы можете как выбрать нужную вам модель и анимацию из большых списков уже готовых моделей и пресетов анимаций, так и добавить свою антропоморфную модель с последующим добавлением скелета и выбором готовой анимации к ней.
Примеры использования:- Быстрое (в 1 клик) назначение скелета к модели. Для этого вам нужно выбрать модель, скелет
- Если вам не подходит конечный результат, то можно выбрать режим “Step-By-Step”
5. Shelf Editor, Shelves и немного о MEL (Maya Embedded Language)
Данная секция содержит в себе общую информацию Shelf Editor, самих shelves, MEL (Maya Embedded Language), Script Editor и о том как начать автоматизировать ваши процессы внутри Maya при помощи MEL.
5.1 Shelf Editor, shelves
Shelves и Shelf Editor крайне полезны и помогают пользователю использовать функциональность Maya в 1 клик. Добавляйте свои shelves, редактируйте уже существующие. В таких элементах интерфейса вы можете добавлять любые инструменты, редакторы и даже сохранять ваши скрипты. Чтобы сделать это вам нужно навести курсор мыши на инструмент, который вы хотите добавить, зажать “Shift + Ctrl” и кликнуть на элемент. После этого он будет добавлен в текущий выбранный shelf.
Примеры использования:
- Быстрое использование комманд, инструментов, редакторов
- Создавайте новые shelves для разных нужд: создание новых объектов, работа с анимацией, все нужные вам редакторы и т.п.
5.2 Maya Embedded Language (MEL)
MEL — это скриптовый язык, лежащий в основе Maya. Пользовательский интерфейс Maya создан преимущественно с использованием MEL. Всё, что вы можете сделать используя UI программы, может быть интерпретировано и автоматизировано при помощи MEL.
Самый простой сценарий для базовой автоматизации чего-либо в Maya — это сделать то, что вам нужно используя UI программы, далее открыть Script Editor (о нём чуть ниже) и сохранить себе команду, которая была использована для выполненной функциональности. Для большего удобства вы можете сохранить данный скрипт в shelf, созданием соответствующей кнопки.
Примеры использования:
- Быстрое использование комманд в обход UI.
- Автоматизация вашей работы
5.3 Script Editor
Script Editor позволяет посмотреть использованные и вводить свои MEL или Python скрипты, сохранять ваши команды в виде кнопки на выбранном shelf, а также просматривать историю использования команд. Открыть Script Editor можно следующими способами:
- В главном меню: Windows -> General Editors -> Script Editor
- При помощи соответствующей кнопки в нижнем левом углу интерфейса Maya
Примеры использования:
- Просматривайте и анализируйте используемые в течении работы команды
- Копируйте, комбинируйте, создавайте и используйте MEL команды для создания своих собственных скриптов
- Сохраняйте скрипты в виде кнопок на ваших shelves для быстрого их использования
На данный момент были перечислены все основные инструменты и редакторы, с которыми я работал наиболее интенсивно. Буду рад вашим комментариям, а также упоминанием той функциональности, которая еще не была перечислена в данной статье!
Новый рекорд рода Yua (Vitaceae) из Вьетнама
Chen, I. (2009) История Vitaceae, выведенная на основе филогении на основе морфологии и летописи окаменелостей семян . Кандидат наук. Тезис. Университет Флориды, Гейнсвилл, 326 стр.
Чен, З.Д., Рен, Х. и Вен, Дж. (2007) Vitaceae. In : Wu, Z.Y., Hong, D.Y. И Рэйвен, П. (Ред.) Флора Китая , 12. Science Press и Missouri Botanical Garden Press, Пекин и Св.Луи, стр. 173–222.
Gilg, E.F. & Diels, L. (1901) Vitaceae. In : Engler, A. (Ed.) Botanische Jahrbücher für Systematik, Pflanzengeschichte und Pflanzengeographie , 29. Schweizerbart, Stuttgart, pp. 460–467.
Jussieu, A.L. de (1818) Columellia . В : Кювье, Г.Ф. (Ред.) Dictionnaire des Sciences Naturelles, dans lequel on traite méthodiquement des différens êtres de la nature, considérés soit en eux-mêmes, d’aprés l’état actuel de nos connoissances, soit relativement à l’utilité re qutireén pe médecine, l’agriculture, le commerce et les arts . изд. 2 10. Ф. Г. Левро, Страсбург, Le Normant, Париж, стр. 102–103.
Лоусон, М.А. (1875) Ampelideae. In : Hooker, J.D. (Ed.) Флора Британской Индии , 1 (3). Л. Рив и Ко, Лондон, стр. 644–668.
Li, C.L. (1990) Yua C.L.Li — новый род Vitaceae. Acta Botanica Yunnanica 12: 1–10.
Li, C.L. (1996) Новые таксоны Vitaceae из Китая. Китайский журнал экологической биологии 2: 43–53.
Линней, К. (1753) Виды Plantarum , 1. Л. Савиус, Стокгольм, 1200 стр.
Лу, Л.М., Ван, В., Чен, З.Д. И Вен Дж. (2013) Филогения немонофилетических Cayratia Juss. (Vitaceae) и значение для эволюции характера и биогеографии. Молекулярная филогенетика и эволюция 68: 502–515.
http://dx.doi.org/10.1016/j.ympev.2013.04.023
Metcalf, F.P. (1948) Критические заметки о Parthenocissus . Бюллетень Института биологии Фан-Мемориал , Ботаника, Пекин 1: 131–137.
Michaux, A. (1803) Flora boreali-americana , 1. Typis Caroli Crapelet, apud Fratres Levrault, Paris & Strasbourg, 330 стр.
Miller, MA, Pfeiffer, W. & Schwartz, T. (2010 ) Создание научного шлюза CIPRES для вывода больших филогенетических деревьев . Новый Орлеан, Лос-Анджелес, 8 стр.
http://dx.doi.org/10.1109/gce.2010.5676129
Нгуен, Х.Х. (2004) Vitaceae. In : Nguyen, T.B. (Ред.) Контрольный список видов растений Вьетнама , 2. Покрытосеменные. Сельскохозяйственное издательство, Ханой, стр. 1145–1160.
Nguyen, T.C. (2012) Таксономическая ревизия семейства виноградных (Vitaceae Juss.) Из Вьетнама . Кандидатская диссертация. Институт экологии и биологических ресурсов Вьетнамской академии наук и технологий, Ханой, 122 стр.
Nylander, J.A.A. (2004) MrModeltest v2 . Программа распространяется автором.Центр эволюционной биологии Упсальского университета.
Pham, H.H. (2003) An Illustrated Flora Vietnam, 2. Young Publishing House, Hochiminh, 951 стр.
Planchon, J.E. (1884) Ampelocissus Planch. Les vignes américaines: leur culture, leur résistance au Phylloxera et leur avenir en Europe , 8 (1): 370–381.
Planchon, J.E. (1887) Monographie des Ampélidées vrais. In: De Candolle, A.F.P.P. и де Кандоль, К.(Ред.) Monographiae Phanaerogamarum , 5. Sumptibus G. Masson, Париж, стр. 305–654.
Reid, E.M. & Chandler, M.E.J. (1933) Лондонская глиняная флора . Британский музей, Лондон, 561 стр.
http://dx.doi.org/10.5962/bhl.title.110147
Ронквист, Ф., Тесленко, М., Марк, П., Эйрес, Д.Л., Дарлинг, A., Höhna, S., Larget, B., Liu, L., Suchard, MA & Huelsenbeck, JP (2012) MrBayes 3.2: эффективный байесовский филогенетический вывод и выбор модели в большом модельном пространстве. Систематическая биология 61: 539–542.
http://dx.doi.org/10.1093/sysbio/sys029
Shetty, B.V. & Singh, P. (2000) Vitaceae. В : Сингх, Н.П., Вохра, Дж. Н., Хаджра, П.К. И Сингх, Д.К. (Ред.) Флора Индии , 5. Ботаническое исследование Индии, Калькутта, стр. 246–324.
Стаматакис А. (2006) RAxML-VI-HPC: филогенетический анализ на основе максимального правдоподобия с тысячами таксонов и смешанными моделями. Биоинформатика 22: 2688–2690.
http://dx.doi.org/10.1093/bioinformatics/btl446
Stuntz, S.C. (1915) 34578. Psedera thomsoni (Lawson) Stuntz. In : Taylor, WA (Ed.) Инвентаризация семян и растений, импортированных, , проведенная Управлением иностранных семян и интродукции растений в период с 1 октября по 31 декабря 1912 г. , № 33. Правительственная типография , Вашингтон, стр. 35.
Wen, J. (2007) Vitaceae. In: Kubitzki, K. (Ed.) Семейства и роды сосудистых растений, 9.Springer-Verlag, Берлин, стр. 466–478.
http://dx.doi.org/10.1007/978-3-540-32219-1_54
Вен, Дж., Лу, Л.М., Не, З.Л., Манчестер, С.Р., Икерт-Бонд, С.М. И Чен, З.Д. (2015) Филогенетика и пересмотренная классификация Vitaceae . Ботаника 2015. Эдмонтон, Альберта.
Zhang, N., Wen, J. & Zimmer, E.A. (2015) Паттерны экспрессии ортологов AP1, FUL, FT и LEAFY у Vitaceae подтверждают гомологию усиков и соцветий во всем семействе винограда. Журнал систематики и эволюции 53: 469–476.
http://dx.doi.org/10.1111/jse.12138
(PDF) Молекулярная филогения и биогеографическая диверсификация Parthenocissus (Vitaceae), разделяющая Азию и Северную Америку
1350 Американский журнал ботаники [Vol. 97
ЛИТЕРАТУРА
Адзума, Х., Дж. Г. Гарк-а-Франко, В. Рико-Грей и Л. Б. Тьен. 2001 г.
Молекулярная филогения магнолиевых: биогеография
разделений тропических и умеренных зон.Американский журнал ботаники 88:
2275 — 2285.
Брижицки, Г. К. 1965. Роды Vitaceae на юго-востоке США
штатов. Журнал Дендрария Арнольда 46: 48 — 67.
Браун Р. У. 1962. Палеоценовая флора Скалистых гор и Великих
равнин. Профессиональный документ геологической службы США 375: 1 — 120.
Чандлер, М. Э. Дж. 1957.Олигоценовая флора озера Бови Трейси
Бассейн, Девоншир. Бюллетень Британского музея (естественная история),
Историческая серия 3: 103 — 106 (Естественная история).
Чандлер, М. Э. Дж. 1961. Нижние третичные флоры южной Англии.
I. Палеоценовые флоры. London Clay ora (приложение), текст и атлас.
Британский музей (естественная история), Лондон, Великобритания.
Чандлер, М.Э. Дж. 1962. Нижние третичные флоры южной Англии.
II. Флора трубочно-глиняной серии Дорсета (Нижний Багшот). Британский
Музей (Естественная история), Лондон, Великобритания.
Чандлер, М. Э. Дж. 1978. Дополнение к нижнетретичным флорам
южной Англии. Специальная статья по третичным исследованиям 4: 1 — 44.
Чен, И. 2009. История Vitaceae определена на основе морфологической филогении
и летописи окаменелостей семян.Университет Флориды,
Гейнсвилл, Флорида, США.
Чен И. и С. Р. Манчестер. 2007 г. Морфология семян современных
и ископаемых Ampelocissus (Vitaceae) и значение для фитогеографии
phy. Американский журнал ботаники 94: 1534 — 1553.
Chen, Z. D., H. Ren, and J. Wen. 2007 г. Vitaceae. В З.-Й. Ву, Д.-Й.
Хонг и П.Х. Рэйвен [ред.], Флора Китая. Science Press и
Missouri Botanical Garden Press, Пекин, Китай и Сент-Луис,
Миссури, США.
Донохью, М. Дж., К. Д. Белл и Дж. Х. Ли. 2001 г. Филогенетические закономерности в географии растений
Северного полушария. Международный журнал растений
Sciences 162: S41 — S52.
Донохью, М. Дж. И С. А. Смит.2004 г. Образцы в совокупности
лесов умеренного пояса в северном полушарии. Философские
Труды Лондонского королевского общества, B, биологические науки
359: 1633 — 1644.
Дойл, Дж. А. и М. Дж. Донохью. 1993 г. Филогения и разнообразие покрытосеменных
. Палеобиология 19: 141 — 167.
Дойл, Дж. Дж. И Дж. Л.Дойл. 1987 г. Процедура быстрого выделения ДНК из
небольших количеств свежих тканей листа. Фитохимический бюллетень 19: 11-15.
Драммонд, А. Дж., С. Ю. У. Хо, М. Дж. Филлипс и А. Рамбо. 2006 г.
Расслабленная филогенетика и доверительные датировки. PLoS Biology 4: e88.
Драммонд, А. Дж., С. Ю. У. Хо, Н. Роленс и А.Рамбаут. 2007 г. Пример
для BEAST 1.4. Оклендский университет, Окленд, Новая Зеландия
. Сайт http://code.google.com/p/beast-mcmc/downloads/list.
Драммонд, А. Дж. И А. Рамбо. 2007 г. BEAST: байесовская эволюция —
анализ с помощью выборки деревьев. BMC Evolutionary Biology 7: 214.
Фаррис, Дж. С., М. Кэллерсйё, А.Г. Клюге и К.Bult. 1994 г. Проверка
на значимость несоответствия. Кладистика 10: 315 — 319.
Фельзенштейн, Дж. 1985. Пределы уверенности в филогенезе: подход, использующий бутстрап. Эволюция 39: 783 — 791.
Фельзенштейн, Дж. 1988. Филогенез из молекулярных последовательностей: вывод
и надежность. Ежегодный обзор генетики 22: 521 — 565.
Fitch, W.М. 1971. К определению хода эволюции: минимум
изменений для определенной топологии дерева. Систематическая зоология 20: 406 — 416.
Галет, П. 1967. Исследует методы идентификации и классификации
температур Vitac é es. II Эти документы присылаются а ля Faculte des Sciences
de Montpellier. Университет Монпелье, Монпелье, Франция.
Гандольфо, М.А., К. К. Никсон и В. Л. Крепет. 2008 г. Отбор
окаменелостей для калибровки моделей молекулярного датирования. Анналы
Ботанический сад Миссури 95: 34 — 42.
Gradstein, F., J. Ogg, and A. B. Smith. 2004 г. Геологическая шкала времени
2004. Cambridge University Press, Кембридж, Великобритания.
Хао Г., Р. М. К. Сондерс и М.L. Chye. 2000 г. Филогенетический анализ Illiciaceae на основе последовательностей внутренних транскрибированных пространств
ers (ITS) ядерной рибосомной ДНК. Систематика и эволюция растений
223: 81 — 90.
Харви Б. Т. 1915. Распространение семян лианы Вирджинии
английских воробьев. Растительный мир 18: 217 — 219.
Huelsenbeck, J. P., and R. Ronquist.2001 г. MrBayes: байесовский вывод
филогенетических деревьев. Биоинформатика 17: 754 — 755.
Икерт-Бонд, С. М. и Дж. Вен. 2006 г. Филогения и биогеография
Altingiaceae: данные комбинированного анализа пяти некодирующих участков хромопласта
. Молекулярная филогенетика и эволюция 39: 512 — 528.
Кирххаймер, Ф. 1938. Beitr ä ge zur n ä heren Kenntnis von Vitaceen-
Samenformen terti ä ren Alters.Планта 28: 582 — 598.
Кирххаймер, Ф. 1939. Rhamnales I: Vitaceae. В W. Jongmans
[ed.], Fossilium catalogus II: Plantae. Dr. W. Junk, Verlag fur
Naturwissenschaften, ‘s-Gravenhage, Нидерланды.
Ли, К. Л. 1990. Юа К.Л. Li — — Новый род Vitaceae. Acta Botanica
Юннаника 12: 1 — 10.
Ли, К. Л. 1998. Vitaceae, Flora Republicae Popularis Sinica, т.48.
Science Press, Пекин, Китай.
Мэддисон, У. П. и Д. Р. Мэддисон. 2009 г. Mesquite: модульная система
для эволюционного анализа, версия 2.72. Веб-сайт http: // mesquit-
eproject.org.
Магальон С. и М. Дж. Сандерсон. 2001 г. Абсолютные коэффициенты диверсификации
в кладах покрытосеменных. Эволюция 55: 1762 — 1780.
Май, Д.Х. 1987 г. Neue Fr ü chte und Samen aus pal ä oz ä nen Ablagerungen
Mitteleuropas. Feddes Repertorium 98: 197 — 229.
Манчестер, С. Р. 1994. Плоды и семена среднего эоцена. Орех
Слои Флора, формация Кларно, Орегон. Palaeontographica Americana
58: 1 — 205.
Манчестер, С. Р., К. Б. Пигг и П. Р. Крейн. 2004 г. Palaeocarpinus
dakotensis sp.п. (Betulaceae: Coryloideae) и связанные с ними тычиночки
серёжки, пыльца и листья из палеоцена Северной Дакоты.
Международный журнал наук о растениях 165: 1135 — 1148.
Мартин А. К., Х. С. Зим и А. Л. Нельсон. 1961 г. Американская дикая природа и
растений: Руководство по пищевым привычкам диких животных. Довер, Минеола, Нью-Йорк,
США.
Моррис, А. Б., К.Д. Белл, Дж. У. Клейтон, В. С. Джадд, Д. Э. Солтис и
П. С. Солтис. 2007 г. Филогения и оценка времени расхождения в
Illicium с последствиями для биогеографии Нового Света. Систематический
Ботаника 32: 236 — 249.
Ниа, Т. Дж. И М. Дж. Сандерсон. 2004 г. Оценка качества оценок времени расхождения молекул —
путем калибровки окаменелостей и выбора модели
на основе окаменелостей.Философские труды Королевского общества
Лондон, Б., Биологические науки 359: 1477 — 1483.
Nie, Z.-L. , H. Sun, Y. Meng и J. Wen. 2009 г. Филогенетический анализ
Toxicodendron (Anacardiaceae) и его биогеографические последствия
для эволюции северных умеренных и тропических межконтинентальных стыков. Журнал систематики и эволюции 47: 416 — 430.
Nie, Z.-L. , J. Wen, H. Azuma, Y.-L. Цю, Х. Сунь, Ю. Мэн, В.-Б. Sun, и
E. A. Zimmer. 2008 г. Филогенетическая и биогеографическая сложность
Magnoliaceae в северном полушарии, определенная на основе трех ядерных наборов данных
. Молекулярная филогенетика и эволюция 48: 1027-1040.
Нюландер, Дж. А. А. 2004.MrModeltest, версия 2 [компьютерная программа].
Центр эволюционной биологии, Уппсальский университет, Упсала, Швеция.
Паркс, К. Р. и Дж. Ф. Вендель. 1990 г. Молекулярная дивергенция между
азиатскими и североамериканскими видами Liriodendron (Magnoliaceae)
с последствиями для интерпретации ископаемых флор. Американский журнал
Ботаники 77: 1243 — 1256.
Планшон, Дж.Э. 1887 г. Monographie des Amp é lid é es vrais. В A. F. P. P. de
Candolle и C. de Candolle [ред.], Monographiae Phanaerogamarum
vol. 5, 305–654. Г. Мейсон, Париж, Франция.
Принс Л. М. и К. Р. Паркс. 2001 г. Филогенетические отношения
Theaceae, выведенные из данных последовательности ДНК хлоропластов. Американский
Ботанический журнал 88: 2309-2320.
Рамбаут, А.2002 г. Se-Al: редактор выравнивания последовательностей, версия 2.0a11.
Зоологический факультет Оксфордского университета, Оксфорд, Великобритания. Веб-сайт
http://tree.bio.ed.ac.uk/software/seal/.
Редер А.А. 1945. Moraceae, Hippocastanaceae et Vitaceae, nomina
conservanda. Журнал Дендрария Арнольда 26: 277 — 279.
Россетто М., Б. Р. Джексы, К. Д. Скотт и Р. Дж. Генри.2001 г. Межродовые отношения
у австралийских Vitaceae: новые данные анализа хпДНК
. Генетические ресурсы и эволюция сельскохозяйственных культур 48: 307 — 314.
SEC.gov | Превышен порог скорости запросов
Чтобы обеспечить равный доступ для всех пользователей, SEC оставляет за собой право ограничивать запросы, исходящие от необъявленных автоматизированных инструментов. Ваш запрос был идентифицирован как часть сети автоматизированных инструментов за пределами допустимой политики и будет обрабатываться до тех пор, пока не будут приняты меры по объявлению вашего трафика.
Укажите свой трафик, обновив свой пользовательский агент, включив в него информацию о компании.
Чтобы узнать о передовых методах эффективной загрузки информации с SEC.gov, в том числе о последних документах EDGAR, посетите sec.gov/developer. Вы также можете подписаться на рассылку обновлений по электронной почте о программе открытых данных SEC, включая передовые методы, которые делают загрузку данных более эффективной, и улучшения SEC.gov, которые могут повлиять на процессы загрузки по сценариям. Для получения дополнительной информации свяжитесь с opendata @ sec.губ.
Для получения дополнительной информации см. Политику конфиденциальности и безопасности веб-сайта SEC. Благодарим вас за интерес к Комиссии по ценным бумагам и биржам США.
Код ссылки: 0.7ecef50.1636225548.2433438b
Дополнительная информация
Политика безопасности в Интернете
Используя этот сайт, вы соглашаетесь на мониторинг и аудит безопасности. В целях безопасности и обеспечения того, чтобы общедоступная служба оставалась доступной для пользователей, эта правительственная компьютерная система использует программы для мониторинга сетевого трафика для выявления несанкционированных попыток загрузки или изменения информации или иного причинения ущерба, включая попытки отказать пользователям в обслуживании.
Несанкционированные попытки загрузить информацию и / или изменить информацию в любой части этого сайта строго запрещены и подлежат судебному преследованию в соответствии с Законом о компьютерном мошенничестве и злоупотреблениях 1986 года и Законом о защите национальной информационной инфраструктуры 1996 года (см. Раздел 18 USC §§ 1001 и 1030).
Чтобы обеспечить хорошую работу нашего веб-сайта для всех пользователей, SEC отслеживает частоту запросов на контент SEC.gov, чтобы гарантировать, что автоматический поиск не влияет на возможность доступа других пользователей к SEC.содержание правительства. Мы оставляем за собой право блокировать IP-адреса, которые отправляют чрезмерное количество запросов. Текущие правила ограничивают пользователей до 10 запросов в секунду, независимо от количества машин, используемых для отправки запросов.
Если пользователь или приложение отправляет более 10 запросов в секунду, дальнейшие запросы с IP-адреса (-ов) могут быть ограничены на короткий период. Как только количество запросов упадет ниже порогового значения на 10 минут, пользователь может возобновить доступ к контенту на SEC.губ. Эта практика SEC предназначена для ограничения чрезмерного автоматического поиска на SEC.gov и не предназначена и не ожидается, чтобы повлиять на людей, просматривающих веб-сайт SEC.gov.
Обратите внимание, что эта политика может измениться, поскольку SEC управляет SEC.gov, чтобы гарантировать, что веб-сайт работает эффективно и остается доступным для всех пользователей.
Примечание: Мы не предлагаем техническую поддержку для разработки или отладки процессов загрузки по сценарию.
Delta Air Lines объявляет 24-й квартал подряд дивидендов
АТЛАНТА, 18 апреля 2019 г. / PRNewswire / — Совет директоров Delta Air Lines (NYSE: DAL) объявил сегодня квартальный дивиденд в размере $ 0.35 за акцию. Дивиденды подлежат выплате зарегистрированным акционерам на момент закрытия рабочего дня 2 мая 2019 г. и будут выплачены 23 мая 2019 г.
Delta Air Lines (NYSE: DAL) — мировой лидер американских авиакомпаний в области продуктов, услуг, инноваций, надежности и качества обслуживания клиентов. Благодаря 80 000 сотрудников по всему миру, Delta продолжает инвестировать миллиарды в своих сотрудников, улучшая качество авиаперелетов и обеспечивая лучшую в отрасли прибыль для акционеров.
- Delta ежегодно обслуживает около 200 миллионов человек, обеспечивая клиентов своей ведущей в отрасли глобальной сетью более чем в 300 пунктов назначения в более чем 50 странах.
- Компания Delta со штаб-квартирой в Атланте предлагает более 5000 ежедневных вылетов и до 15000 аффилированных вылетов, включая главный альянс SkyTeam, членом-учредителем которого является Delta.
- Благодаря инновационным альянсам с Aeromexico, Air France-KLM, Alitalia, China Eastern, GOL, Korean Air, Virgin Atlantic, Virgin Australia и WestJet, Delta предоставляет клиентам по всему миру больше выбора и конкуренции.
- Delta управляет крупными транспортными узлами и ключевыми рынками в аэропортах Амстердама, Атланты, Бостона, Детройта, Лондон-Хитроу, Лос-Анджелеса, Мехико, Миннеаполиса / Сент.Поль, Нью-Йорк-Джона Кеннеди и Ла-Гуардия, Париж-Шарль-де-Голль, Солт-Лейк-Сити, Сан-Паулу, Сиэтл, Сеул-Инчхон и Токио-Нарита.
- Delta вошла в список 50 самых уважаемых компаний по версии журнала Fortune, а также была названа самой уважаемой авиакомпанией в восьмой раз за девять лет. Кроме того, Delta занимает первое место в ежегодном опросе авиакомпаний Business Travel News в течение беспрецедентных восьми лет подряд и два года подряд признается одной из самых инновационных компаний Fast Company в мире.
- Как работодатель, Delta регулярно удостаивается высших наград от таких организаций, как Glassdoor, и признается лучшим местом работы для женщин и военнослужащих. Генеральный директор Delta Эд Бастиан был назван журналом Fortune одним из «величайших лидеров мира» в 2018 году.
- Более подробную информацию о Delta можно найти в Delta News Hub, а также на сайте delta.com, через @DeltaNewsHub в Twitter и Facebook.com/delta.
ИСТОЧНИК Delta Air Lines
Ссылки по теме
http: // www.delta.com
Наблюдая за рождением айсберга
ПУНТА-АРЕНА, ЧИЛИ — После обнаружения трещины, которая пересекает плавучий шельфовый ледник на леднике Пайн-Айленд в Антарктиде, сотрудники NASA Operation IceBridge выполнили следующую миссию и впервые в истории провели подробные измерения с воздуха крупного айсберга. прогресс.
Операция НАСА «Ледяной мост», крупнейшее из когда-либо совершавшихся аэрофотосъемок полярных льдов Земли, находится в разгаре своей третьей полевой кампании из Пунта-Аренаса, Чили.Шестилетняя миссия даст беспрецедентный трехмерный вид арктических и антарктических ледниковых щитов, шельфовых ледников и морского льда.
Ледник Пайн-Айленд в последний раз откололся значительный айсберг в 2001 году, и некоторые ученые недавно предположили, что он был подготовлен к повторному отелу. Но до полета на IceBridge 14 октября космического корабля НАСА DC-8 никто не видел никаких свидетельств того, что шельфовый ледник начал распадаться. С тех пор более подробный взгляд на спутниковые снимки, кажется, показывает первые признаки трещины в начале октября.
В то время как Пайн-Айленд привлекает внимание ученых, потому что он большой и нестабильный — ученые называют это крупнейшим источником неопределенности в прогнозах глобального повышения уровня моря, — происходящий сейчас отел является частью естественного процесса для ледника, который заканчивается в открытой воде. . Гравитация тянет лед в леднике на запад вдоль Гудзоновых гор в Антарктиде к морю Амундсена. Плавающий ледяной язычок простирается на 30 миль в Амундсен за линией заземления, точкой ниже уровня моря, где шельфовый ледник замыкается на континентальной скале.По мере того как лед продвигается к морю изнутри, шельфовый ледник неизбежно треснет и выбросит на свободу большой айсберг.
«На самом деле сейчас мы наблюдаем, как это происходит, и это очень волнует для нас», — сказал ученый проекта IceBridge Майкл Студингер, Центр космических полетов Годдарда, Гринбелт, штат Мэриленд. «Это часть естественного процесса, но находиться здесь очень интересно и Насколько мне известно, никто не управлял лидаром над активно развивающимся рифтом, таким как этот.»
Основная цель операции IceBridge — установить одни и те же инструменты на одни и те же линии полета и пути спутников год за годом, чтобы собрать значимые и точные данные о том, как ледяные щиты и ледники меняются с течением времени. Но обнаружение развивающихся трещина в одной из самых важных научных целей в мире гляциологии предложила краткое изменение программы полета 26 октября, хотя бы для 30-минутного отклонения от предписанных дневных линий полета.
Команда IceBridge наблюдала трещину пробегает шельфовый ледник примерно на 18 миль.Лидарный прибор DC-8, Airborne Topographic Mapper, измерил плечи разлома на расстоянии около 820 футов (250 метров) в самом широком месте, хотя разлом простирался примерно на 260 футов вдоль большей части трещины. Самые глубокие точки от поверхности шельфового ледника находились в диапазоне от 165 до 195 футов (от 50 до 60 метров). Когда айсберг выйдет на свободу, он покроет площадь около 340 квадратных миль (880 квадратных километров). Радиолокационные измерения показали, что шельфовый ледник в районе разлома имеет толщину около 1640 футов (500 метров) футов, из которых только около 160 футов плавает над водой, а остальные находятся под водой.Вполне вероятно, что после того, как айсберг уплывет, передняя кромка шельфового ледника отступит дальше, чем когда-либо с тех пор, как его местоположение было впервые зарегистрировано в 1940-х годах.
| В октябре 2011 года в рамках операции «Ледяной мост» НАСА обнаружила крупный разрыв в леднике Пайн-Айленд в западной Антарктиде. Эта трещина, которая простирается не менее 18 миль и имеет глубину 50 метров, может образовать айсберг размером более 800 квадратных километров. Ученые IceBridge вернулись вскоре после этого, чтобы провести первые в истории подробные воздушные измерения крупного айсберга, который только что начал откалываться.Предоставлено: НАСА / Годдард / Джефферсон Бек, | .
Пилот-ветеран DC-8 Билл Брокетт первым выполнил запланированную на день миссию, пересекая поток ледника возле линии заземления, чтобы собрать данные о его высоте, топографии и толщине. Когда пришло время исследовать трещину, Брокетт перелетел через нее, а затем повернулся и полетел вдоль трещины на виду. Банкомат составляет точные топографические карты с помощью лазера, который сканирует на 360 градусов 20 раз в секунду, при этом генерируя 3000 лазерных импульсов в секунду.При полете на высоте 3000 футов, как и во время этого полета, он измеряет полосу поверхности около 1500 футов шириной. Поскольку ширина трещины местами превышала 800 футов, для Брокетта было важно крепко удерживать ее.
«Пилоты проделали действительно хорошую работу по удержанию самолета и нашей полосы сканирования ATM в значительной степени над разломом, когда вы летели из одного конца в другой», — сказал Джим Юнгель, который возглавляет группу ATM с острова Уоллопс. Летный комплекс в Вирджинии.«Это было настоящим испытанием, когда нам сказали… мы собираемся попытаться пролететь по нему, и давайте посмотрим, могут ли ваши лидарные системы отобразить эту трещину и отобразить ее дно.
» И это было очень весело на личном уровне, чтобы увидеть, действительно ли то, что вы создали за эти годы, может выполнять такую работу. Так что да, мне понравилось. Мне очень понравилось наблюдать за полученными результатами ».
В то время как банкомат обеспечивал наиболее подробные измерения топографии разлома, другие инструменты на борту DC-8 также фиксировали уникальные аспекты.Цифровая картографическая система, камера обзора надира, собрала крупные планы высокого разрешения скалистого ущелья. Во время полета перпендикулярно трещине радар McCORDS также измерил ее глубину и толщину шельфового ледника в этом районе.
Чтобы поймать трещину в действии, требовалось немного удачи, но это также свидетельство научной пользы от последовательных, повторяющихся рейсов и гибкости пилотируемой миссии в полевых условиях.
«Много раз, когда вы занимаетесь наукой, у вас нет возможности поймать большие истории, поскольку они происходят, потому что вы находитесь не в нужном месте в нужное время», — сказал Джон Зоннтаг, Instrument Руководитель группы операции «Ледяной мост», базируется в Центре космических полетов Годдарда.«Но на этот раз мы были».
Ссылки по теме
›Анимация изменений Антарктики
› Изображение PIG
›Дополнительные изображения IceBridge
› Страница миссии IceBridge
границ | Ризосферные почвенные бактериальные сообщества устойчивых к непрерывному возделыванию и чувствительных генотипов сои по-разному реагируют на длительное непрерывное возделывание моллисолей
Введение
Являясь одним из важнейших почвенных ресурсов Китая, моллисоли на северо-востоке страны играют решающую роль в поддержании внутреннего спроса на продовольствие.Соя [ Glycine max (L.) Merrill] — одна из самых важных продовольственных культур в мире, обеспечивающая большое количество масла и белка для людей и животных (Chigen et al., 2018; Liu et al., 2020 ). Соевые бобы пользуются большим спросом, особенно в Китае (Liu et al., 2020). Из-за ограниченности пахотных земель, климатических условий и большой доли пахотных земель с другими культурами в этом регионе часто непрерывно выращивают сою (Liu et al., 2012). На некоторых фермах соя выращивалась непрерывно в течение 40 лет или даже дольше (Liu et al., 2020). Непрерывный посев сои приводит к закислению почвы, обострению заболеваний, передаваемых через почву, снижению активности ферментов почвы и накоплению токсинов в почве (Zhan et al., 2004; Yan et al., 2012; Pérez-Brandán et al. ., 2014; Бай и др., 2015). Несколько исследований показали, что эти изменения в значительной степени связаны с биологическими факторами почвы (Ji et al., 1996; Dias et al., 2015; Liu et al., 2020).
Микроорганизмы ризосферы играют очень важную роль в поддержании устойчивости почвенной экосистемы и здоровья растений (Waldrop et al., 2000; Авидано и др., 2005). Различные системы земледелия могут существенно изменить структуру микробного сообщества (Meriles et al., 2009; Zhou et al., 2018; Liu et al., 2019). Однако эти изменения зависят от типа системы земледелия, типа почвы и вида сельскохозяйственных культур. Например, как Tang et al. (2009) и Zhu et al. (2014) обнаружили, что по сравнению с системами севооборота численность актинобактерий значительно снизилась при непрерывном возделывании сои. Xu et al. (1995) обнаружили, что постоянное земледелие снижает численность Penicillium sp., увеличивая численность Fusarium sp. Однако другое исследование показало, что по сравнению с севооборотом непрерывный посев сои не изменил структуру микробного сообщества (Hu and Wang, 1996). Несоответствия между этими исследованиями в основном связаны с различиями в типах почв, методах исследований, системах севооборота и годах непрерывного земледелия. В то же время механизмы, связанные с постоянными препятствиями при возделывании сельскохозяйственных культур, сложны и требуют более глубокого изучения в различных условиях.
Соя — одна из наиболее чувствительных культур к непрерывному земледелию. Исследования показали, что непрерывное выращивание сои в течение 3 лет может снизить урожайность примерно на 30% (Liu et al., 2020). Кроме того, по сравнению с севооборотом, количество заболеваний корневой гнилью сои и цистовых нематод на соевых полях значительно увеличилось в краткосрочной системе непрерывного возделывания культур (Cai et al., 2015). Однако корневая гниль и болезнь кистозных нематод на соевых полях могут быть ослаблены после длительного непрерывного сбора урожая (Song et al., 2017). Это может быть связано с тем, что длительное непрерывное земледелие может увеличить популяцию полезных микроорганизмов, что может помочь подавить колонизацию и развитие патогенов в почве (Wei et al., 2015; Liu et al., 2020). Например, Liu et al. (2020) обнаружили, что 13 лет непрерывного выращивания сои в течение 13 лет увеличивают количество полезных бактерий, таких как Bradyrhizobium sp., Gemmatimonas sp., Mortierella sp. И Paecilomyces sp.и уменьшил количество патогенных грибов Fusarium sp. Однако некоторые исследователи также обнаружили, что длительное непрерывное употребление сои может уменьшить популяции полезных микроорганизмов, таких как Trichoderma , Colloids и Pseudomonas fluorescens (Pérez-Brandán et al., 2014). Следовательно, изменение микробного сообщества, вызванное непрерывным или чередованием посевов, имеет решающее значение для выявления механизмов, лежащих в основе препятствий для непрерывного выращивания сои.Кроме того, непоследовательность этих исследований указывает на необходимость проведения дополнительных исследований микробного сообщества почвы.
На микробное сообщество ризосферы сельскохозяйственных культур также влияют генотипы (Kaisermann et al., 2017; Lian et al., 2019). Устойчивые генотипы могут привлекать некоторые полезные микроорганизмы, чтобы помочь хозяину противостоять различным стрессам (Lebeis et al., 2015; Kwak et al., 2018). Например, генотипы томатов, устойчивые к увяданию Fusarium , могут задействовать большое количество Flavobacterium для облегчения симптомов увядания Fusarium (Kwak et al., 2018). Наши предыдущие исследования также показали, что устойчивые к алюминию соевые бобы могут привлекать Tumebacillus и Burkholderia и снижать токсичность алюминия (Lian et al., 2019). Что касается непрерывного выращивания сои, хотя известно, что микроорганизмы играют очень важную роль в смягчении или усугублении препятствий для непрерывного возделывания культур, сравнение микробного сообщества в ризосфере между различными генотипами не проводилось. Понимание характеристик микробного сообщества ризосферы соевых бобов с непрерывно устойчивым генотипом может помочь нам понять, как эти устойчивые генотипы могут лучше адаптироваться к непрерывному земледелию.
Здесь мы стремились охарактеризовать ризосферные микроорганизмы разных генотипов сои, один разводится для непрерывного возделывания, а другой нет, выращиваемый в моллизоле на северо-востоке Китая при непрерывном возделывании в течение 7 и 36 лет с севооборотом соя-кукуруза. Основываясь на более высокой адаптивности генотипов сои, устойчивых к непрерывному возделыванию (CC-T), к непрерывному возделыванию, мы предположили, что (1) бактериальное разнообразие в ризосфере генотипов CC-T будет выше, чем у генотипов, устойчивых к постоянному возделыванию (CC -S) генотипов и что (2) реакция структуры бактериального сообщества на непрерывное выращивание между генотипами сои CC-T и CC-S будет разной.CC-T может привлекать полезные виды микробов, чтобы уменьшить распространение болезней, передаваемых через почву.
Материалы и методы
Источники почвы и растительные материалы
Для оценки реакции устойчивых к выращиванию и чувствительных к сельскохозяйственным культурам генотипов сои в данном исследовании использовалась почва Mollisol в соответствии с таксономией почв Министерства сельского хозяйства США. Соевые бобы, использованные в эксперименте, включали Qinong1 и Qinong5, которые были устойчивы к непрерывному выращиванию (CCT), и Heihe43 и Henong76, которые были чувствительны к постоянному выращиванию (CCS).
Опытный образец и коллекция почв растений и ризосферы
Для этого исследования было выбрано непрерывное выращивание двух генотипов сои в течение 7 и 36 лет, а также обработка севооборотов соя-кукуруза в течение 10 лет. Общая площадь экспериментального поля (47 ° 15 ′ с.ш., 123 ° 40 ′ в.д.) составляла 2 га, в том числе 1,3 га для непрерывной 36-летней посевной культуры и по 0,3 га для 7-летней непрерывной и ротационной обработки (дополнительный рисунок S1). Всего на стадии цветения сои 15 июля 2020 г. было отобрано 72 образца почвы ризосферы (шесть обработок × 12 повторностей).Все образцы почвы ризосферы собирали, осторожно встряхивая корень растения, чтобы удалить неплотно прикрепленную почву, а затем почву, приставшую к корневой системе, переносили в асептический мешок. Два грамма ризосферной почвы из каждого образца помещали в стерилизованную микроцентрифужную пробирку и хранили при -80 ° C для экстракции ДНК. Побеги и корни были разделены для измерения биомассы. Все нематоды цист сои ( Heterodera glycines ) и клубеньки из корней были подсчитаны после удаления из корневой системы.Вкратце, после того, как рост нематод цист сои прорвался через поверхность корней сои (через 35 дней после всхода), для подсчета нематод цист сои случайным образом были выбраны 10 растений сои. Урожайность с квадратного метра измеряли после созревания сои.
Извлечение ДНК из почвы и секвенирование нового поколения
Всего для секвенирования было использовано 72 образца почвы ризосферы. Общую ДНК почвы экстрагировали с использованием набора Fast DNA SPIN для почвы (MP Biomedicals, Санта-Ана, Калифорния).Последовательности праймеров 341F / 805R использовали для амплификации гипервариабельных областей V4 (V3 – V4) 16S рРНК. Продукты ПЦР очищали и затем секвенировали с использованием платформы MiSeq Illumina (Illumina, США) в компании Icongene (Wuhan) Gene Technology Co., Ltd. (Ухань, Китай). Все последовательности были депонированы в архив короткого чтения GenBank PRJNA732989.
После секвенирования необработанные файлы FASTQ обрабатывались с помощью конвейера QIIME версии 1.19.1. Вкратце, все считывания последовательности были назначены каждому образцу на основе штрих-кодов.Последовательности с низким качеством (длина <200 пар оснований и средний показатель качества основания <20) были удалены перед дальнейшим анализом. Химера обрезанных последовательностей была обнаружена и удалена с использованием алгоритма UCHIME (Edgar et al., 2011). Последовательности были филогенетически назначены в соответствии с их наилучшим соответствием последовательностям в базе данных RDP с использованием классификатора RDP (Cole et al., 2009). Операционные таксономические единицы (OTU) были классифицированы по сходству последовательностей 97% с использованием CD-HIT (Li and Godzik, 2015). Богатство Chao1 и индекс разнообразия Шеннона были рассчитаны в QIIME.Анализ ограниченных главных координат (CPCoA) и тесты значимости (тест Адониса и тест Mantel) были выполнены в программе R версии 3.5.1 для Windows с «веганским» пакетом. Были построены тройные графики, чтобы показать сравнение численности OTU (> 5%) для системы севооборота и непрерывного выращивания в течение 7 и 36 лет сои двух генотипов с использованием пакета «vcd» (Friendly and Meyer, 2015). Односторонний дисперсионный анализ (ANOVA) использовался в GenStat 13 (VSN International, Hemel Hempstead, Великобритания) для оценки относительной численности бактерий на различных таксономических уровнях.Кроме того, был проведен двумерный корреляционный анализ Пирсона, чтобы получить доступ к корреляциям между pH, нематодами цист сои, урожайностью и родами с более высокой относительной численностью.
Базовые бактерии и анализ сети совместного возникновения
Core бактерий, которые содержат список ОТЕ, наблюдаемых в 60% всех образцов ризосферы, были получены MicrobiomeAnalyst (Chong et al., 2020). Сети бактериальной совместимости были проанализированы для каждой системы земледелия. Чтобы изучить сетевую структуру OTU с высокой численностью, мы выбрали OTU с более чем 0.Относительная численность 2% для расчета коэффициентов ранговой корреляции Спирмена. Корреляции между OTU были выбраны при p <0,05 и коэффициенте корреляции Спирмена более 0,8 (Mendes et al., 2018). Узлы и края представляют бактериальные OTU и корреляции между бактериальными OTU, соответственно. Статистический анализ был рассчитан с использованием пакета «псих» в R, а затем визуализирован в Gephi (Jiang et al., 2017). Виды краеугольного камня были определены в соответствии с высокой степенью узлов, высокой промежуточной центральностью и высокой степенью близости центральности (Berry and Widder, 2014; Agler et al., 2016). Анализ NetShift был проведен с использованием программного обеспечения NetShift для сравнения сетей различных обработок и поиска «микробов-драйверов» между обработкой севооборотов соя-кукуруза и 7,36-летним непрерывным выращиванием урожая для чувствительных и толерантных генотипов, соответственно (Kuntal et al. ., 2019).
Результаты
Количество и урожайность нематод кисты сои
Как системы возделывания, так и генотипы сои значительно повлияли на количество и урожайность цистовых нематод сои (Таблица 1).Число цистовых нематод в чувствительных генотипах было выше, чем в толерантных генотипах. В частности, для чувствительных генотипов количество нематод соевых бобов значительно увеличилось на 7-летнем поле непрерывного возделывания, но заметно снизилось на 36-летнем непрерывном посевном поле. Однако непрерывное земледелие не оказало значительного влияния на толерантные генотипы. Кроме того, урожайность увеличилась на 7-летнем поле непрерывного возделывания, но снизилась на 36-летнем непрерывном посевном поле, с более высокой урожайностью для толерантных генотипов, чем для чувствительных генотипов (Таблица 1).
Таблица 1 . PH почвы, количество цистовых нематод, биомасса побегов и корней, а также урожайность различных обработок.
Бактериальное разнообразие ризосферы почвы
Как системы выращивания, так и генотипы сои не оказали значительного влияния на альфа-разнообразие сообществ ризобактерий (односторонний дисперсионный анализ, p > 0,05; рисунки 1A, B). Что касается бета-разнообразия, CPCoA и двусторонний PERMANOVA показали, что системы возделывания и генотипы сои интерактивно влияют на сообщества ризобактерий (PERMANOVA, p <0.05; Фигура 2; Дополнительный рисунок S2; Таблица 2). Более подробно, структуры сообществ ризобактерий различались между толерантными и чувствительными генотипами сои независимо от систем возделывания (дополнительный рисунок S3; таблица 2).
Рисунок 1 . Ризобактериальное сообщество индекс Шеннона (A) и богатство Chao1 (B) различных обработок. Односторонний дисперсионный анализ, n = 12, p <0,05.
Рисунок 2 . Анализ ограниченных главных координат (CPCoA), основанный на различиях Брея-Кертиса в разнообразии 16S рРНК в ризосфере.
Таблица 2 . Влияние систем возделывания и генотипов сои на дифференциацию бактериальных сообществ по данным PERMANOVA.
Специфические микробиомы двух генотипов сои в различных системах возделывания
Proteobacteria, Bacteroidetes, Firmicutes, Acidobacteria, Actinobacteria и Verrucomicrobia были самыми многочисленными бактериальными типами, живущими в ризосфере при всех обработках, составляя 80,57–82,98% всего сообщества (Рисунок 3; Дополнительная таблица S1).Анализ численности филумов с помощью двухфакторного дисперсионного анализа (дополнительная таблица S1) показал, что системы земледелия значительно ( p <0,05) влияли на 10, 7 и 10 типов. Кроме того, на них значительно ( p <0,05) влияли системы возделывания, генотипы сои и их взаимодействие (дополнительная таблица S1). Более того, реакция на продолжающееся выращивание у двух генотипов была разной. Например, относительная численность ацидобактерий была значительно увеличена на 7-летнем поле непрерывного возделывания, но снизилась относительная численность обоих генотипов сои на 36-летнем непрерывном посевном поле, и с более высокой относительной численностью для чувствительных генотипов, чем для толерантных генотипов. .Относительная численность Proteobacteria значительно снизилась, а затем увеличилась в этих системах возделывания для толерантных генотипов, в то время как она постоянно снижалась для чувствительных генотипов. На уровне рода 16, 20 и 22 рода были значительно ( p <0,05) под влиянием систем возделывания, генотипов сои и их взаимодействия, соответственно (Таблица 3). Среди них некоторые роды разных генотипов по-разному реагируют на непрерывное земледелие. Например, количество Pseudomonas , принадлежащих к Proteobacteria, в ризосфере толерантного генотипа увеличивалось, а затем уменьшалось с непрерывным временем выращивания, тогда как в чувствительных генотипах наблюдалась противоположная тенденция.Однако есть также некоторые роды, которые реагируют на непрерывное выращивание в рамках постоянной тенденции между двумя генотипами. Например, Streptomyces и Bacillus , которые принадлежат к Actinobacteria и Firmicutes, соответственно, снизились на 7-летнем непрерывном поле посевов, но увеличились на 36-летнем непрерывном поле посевов с более высокой относительной численностью в чувствительных генотипах. Анализ линейной модели был использован для идентификации бактериальных ОТЕ, значительно обогащенных толерантными и чувствительными генотипами ризосферной почвы в системе севооборота, 7-летней системе непрерывного земледелия и 36-летней системе непрерывного земледелия.Что касается толерантных генотипов, OTU, принадлежащие к Chitinophaga , Nitrospira и Saprospirales , доминировали в ризосферах системы севооборота, 7-летней системы непрерывного земледелия и 36-летней системы непрерывного земледелия, соответственно (Рисунок 4A). Что касается чувствительных генотипов, ОТЕ, принадлежащие к Phyllobacteriaceae , Comamonadaceae и Cytophagaceae , доминировали в ризосферах системы севооборота, 7-летней системы непрерывного земледелия и 36-летней системы непрерывного земледелия, соответственно (Рисунок 4B).Более подробная информация доступна в дополнительных файлах (дополнительная таблица S2).
Рисунок 3 . Относительное обилие филюмов разных обработок.
Таблица 3 . Системы возделывания, генотипы сои и их интерактивное влияние на относительную численность бактерий на уровне рода.
Рисунок 4 . Тройные графики, изображающие компартменты, показывающие распределение дифференциации сообществ между различными системами земледелия для толерантных генотипов (A) и чувствительных генотипов (B) .Положение каждого круга зависит от его вклада в общую относительную численность. Цветными кружками обозначены OTU, которые обогащены в различных системах земледелия, а зеленые кружки — для системы севооборота; красный и зеленый для систем 7- и 36-летнего непрерывного земледелия соответственно. Серые круги представляют собой OTU, которые существенно не обогащены в заданном диапазоне.
Корреляция между ризосферной почвенной микробиотой с более высокой относительной численностью и pH, нематодами соевых бобов и урожайностью была получена с помощью корреляционного анализа Пирсона (дополнительный рисунок S4).PH почвы положительно коррелировал с Acinetobacter и Bradyrhizobium , тогда как он отрицательно коррелировал с другими родами. Цистовая нематода отрицательно коррелировала с Thermomonas , Flavisolibacter и Opitutus , в то время как положительно коррелировала с Pseudomonas, Renibacterium , Agrobacterium , Bacillus и Streptococcus . Более того, урожай положительно коррелировал с Sphingobium , Flavisolibacter и Opitutus .
Основной микробиом и сеть совместного возникновения
Среди 12 436OTU мы обнаружили, что толерантные и чувствительные генотипы имеют почти одинаковые основные микроорганизмы, такие как OTU3795 ( Thermomonas ), OTU6911 ( norank_Sphingomonadaceae ), OTU8585 ( norank _ Sphing Erythrobacteraceae ), OTU9116 ( norank_Methylophilaceae ), OTU9219 ( Agrobacterium, ), OTU4334 ( Nitrospira ), OTU6274 ( norank_Chitinophagaceae ), OTU6533 ( Flavisolibacter ), OTU10447 ( Renibacterium ), OTU4280 ( norank_ii1-15 ), OTU4827 ( norank_Ellin6075 ), OTU8697 ( norank_RB41 ) и OTU10068 ( norank_ii1-15 ; Рисунок 5).Более того, OTU7122 ( Pseudomonas ) был основным видом, специфичным для чувствительных генотипов (рис. 5B). Относительная численность общих основных OTU толерантных и чувствительных протеобактерий составляла 7,77% и 11,48, соответственно (дополнительная таблица S3).
Рисунок 5 . Относительная численность основного микробиома для толерантного (A) и чувствительного (B) генотипов.
Используя объединенные данные 16S рРНК двух генотипов сои, выращенных в разных системах возделывания, сеть совместного присутствия в ризосфере показала заметные различия между обработками (рис. 6; дополнительная таблица S4).Для толерантного генотипа количество отрицательных корреляций и модульности (M) увеличивалось с годами непрерывного выращивания, что позволяет предположить, что между доминантными бактериальными родами происходило больше событий сцепления, сотрудничества и обмена. Более того, не было значительного изменения среднего коэффициента кластеризации (avgCC) или средней степени (avgK). Для чувствительного генотипа модульность (M) увеличивалась с увеличением продолжительности выращивания сельскохозяйственных культур. Однако количество отрицательных корреляций, средний коэффициент кластеризации (avgCC) и средняя степень (avgK) увеличились в области 7-летних систем непрерывного земледелия, но уменьшились в области 36-летних систем непрерывного земледелия.Ключевые виды бактериальной сети в ризосфере были идентифицированы путем вычисления степени узла, центральности близости и центральности промежуточности для всех узлов в сети (дополнительная таблица S5). В общем, OTU11321 ( Luteolibacter ), OTU6533 ( Flavisolibacter ), OTU1003 ( Chitinophagaceae ), OTU4362 ( norank_RB41 ), OTU4362 ( norank_RB41 ), OTU6549 (113) были идентифицированы как виды Balneon и Balneon ), OTU6575 для двух генотипов, выращенных в разных системах земледелия.Используя анализ NetShift, общий график подсети показал, что «микробами-драйверами» были OTU2820 ( Неклассифицированные Chitinophagaceae ), OTU881 ( Неклассифицированные oc28 ) и OTU3463 ( Неклассифицированные Acidobacteria-6 чередований) для сети. система непрерывного посева и OTU881 ( неклассифицированный oc28 ), OTU3487 ( неклассифицированный Gemm-1 ), OTU9219 ( Agrobacterium ) и OTU3461 ( неклассифицированный Ellin6075 ) для системы непрерывного посева на 7 лет между севооборотами.Для чувствительного генотипа сои «микробами-драйверами» были OTU2166 ( Unclassified Piscirickettsiaceae ) и OTU9044 ( Unclassified Erythrobacteraceae ) для сети между севооборотами и 7-летней системой непрерывного возделывания сельскохозяйственных культур, и OTU144813 Неклассифицированные Oxalobacteraceae ), OTU4607 ( Неклассифицированные Cytophagaceae ), OTU3463 ( Неклассифицированные Acidobacteria-6 ) и OTU3416 ( Неклассифицированные OPB35 ) для сети между 7-летними севооборотами и 7-летними севооборотами.
Рисунок 6 . Сетевой анализ бактериального сообщества ризосферы от различных обработок. Узлы представляют собой OTU, закодированные по типам, и масштабируются по количеству соединений (степени узла). Связи нанесены на график при r > 0,8 (положительная корреляция, красный цвет) или r <-0,8 (отрицательная корреляция, синий цвет) и p <0,05.
Рисунок 7 . Общий вид подсети с выделенным узлом «драйвер». Всем узлам, принадлежащим к одному сообществу, случайным образом был присвоен одинаковый цвет.Серые узлы OTU представляют собой узлы, которые присутствуют в обоих случаях, но напрямую взаимодействуют с общей подсетью либо в случае, либо в контрольной группе. Размер узлов был пропорционален их баллам NESH, и узел окрашивался в красный цвет, если расстояние между контрольной и исследуемой группами увеличивалось. Большие и красные узлы являются особенно важными «драйверами». CK VS 7 (36; толерантный или чувствительный), сетевое сравнение севооборотов соя-кукуруза и 7 (36) лет непрерывного возделывания для толерантного или чувствительного генотипа.
Обсуждение
Наши первоначальные гипотезы заключались в том, что соя с толерантным генотипом будет иметь более высокое бактериальное разнообразие ризосферы, чем соя с чувствительным генотипом, а толерантная соя будет рекрутировать микроорганизмы, которые способствовали бы устойчивой устойчивости сельскохозяйственных культур. Наши результаты показали, что не было значительных различий в микробном разнообразии между двумя генотипами сои во всех системах возделывания. Кроме того, была проверена наша вторая гипотеза: толерантные генотипы привлекают определенные микроорганизмы, которые могут помочь сое смягчить заболевания, передаваемые через почву.Короткий период непрерывного возделывания культур неблагоприятен для сои, но длительные периоды непрерывного возделывания смягчат этот недостаток. Более того, разные генотипы сои по-разному реагируют на системы непрерывного возделывания культур, и микроорганизмы, пополняемые толерантными соевыми бобами, могут играть важную роль в смягчении неблагоприятных последствий непрерывного возделывания культур.
В настоящем исследовании мы обнаружили, что не было значительных различий в разнообразии бактериального сообщества при лечении.Предыдущие исследования показали, что увеличение количества культур может привести к увеличению микробного разнообразия почвы. Лю и др. (2020) обнаружили, что бактериальное разнообразие ризосферы снижалось при непрерывном возделывании сои по сравнению с системами севооборота сои-кукурузы. Кроме того, Zhu et al. (2017) и Лю и др. (2020) обнаружили, что бактериальное разнообразие почвы увеличивается с увеличением количества лет непрерывного земледелия. Однако Ли и др. (2010) не обнаружили значительных различий в микробном разнообразии почвы между почвами непрерывного севооборота сои и сои-кукурузы.Это может быть связано с разным pH почвы в этих исследованиях, как Liu et al. проводили свои исследования на кислой почве, в то время как наше исследование проводилось на нейтральной почве. Изменения pH почвы и других физических и химических свойств были в значительной степени связаны с микробным разнообразием, и это изменение было связано с изменениями в секреции корней, таких как органические кислоты, фенолы и флавоноиды, на которые значительно влияли системы земледелия (Venter et al. ., 2016; Liu et al., 2020). Что касается бета-разнообразия, мы обнаружили, что системы посевов и генотипы сои были основными факторами, которые изменили бактериальные сообщества (тесты Adonis, p <0.05). Этот результат согласуется с результатами, полученными Zhu et al. (2017) и Lian et al. (2019), которые указали, что произошли значительные изменения в бактериальных сообществах почвы в системах долгосрочного и краткосрочного непрерывного возделывания сои и в различных генотипах сои.
Относительная численность Proteobacteria была значительно снижена в ризосферных почвах поля системы непрерывного земледелия по сравнению с таковыми в системе севооборота, что указывает на то, что эти бактерии были уменьшены при низком уровне доступных питательных веществ (Fierer et al., 2007; Ли и др., 2014). Относительная численность Pseudomonas , Streptomyces и Bacillus снизилась на 7-летнем поле непрерывного возделывания, но увеличилась на 36-летнем непрерывном посевном поле с более высокой относительной численностью в чувствительных генотипах. Pseudomonas может улучшить солюбилизацию фиксированного фосфора почвы и внесенных фосфатов, что приведет к повышению урожайности сельскохозяйственных культур (Nautiyal, 1999). Более того, метаболиты Pseudomonas могут увеличивать секрецию аминокислот в корнях люцерны, кукурузы и пшеницы (Phillips et al., 2004). Streptomyces может помочь растениям противостоять таким заболеваниям, как увядание и корневая гниль (Lian et al., 2019; Shi et al., 2020). Кроме того, некоторые бактерии, такие как Thermomonas , Flavisolibacter и Opitutus , были отрицательно связаны с нематодами, что позволяет предположить, что эти бактерии могут ингибировать нематод соевых кист. Таким образом, основываясь на функциях упомянутых выше видов, мы предположили, что изменение относительной численности этих бактерий может быть связано с антагонистической активностью этих таксонов против патогенов растений и улучшением содержания питательных веществ в почве.
Мы использовали дифференциальный анализ численности ОТЕ в обработках и наблюдали высокую относительную численность ОТЕ, обогащенных в системах непрерывного земледелия толерантных генотипов, которые принадлежали Chitinophagaceae , Pseudoxanthomonas , Nitrospira и Streptomyces (Рисунок 4; Таблица 4; Таблица 4; 3). Этот результат согласуется с нашей гипотезой о том, что толерантные генотипы задействовали некоторые микроорганизмы, которые играют важную роль в устойчивости соевых бобов к длительному непрерывному выращиванию.OTU4434 ( Nitrospira ), OTU751 ( Pseudoxanthomonas ) и OTU10218 ( Streptomyces ) показали более высокую относительную численность в системе непрерывного земледелия, чем в системе чередования в настоящем исследовании (Рисунок 4; Таблица 3). Во многих исследованиях сообщается, что Nitrospira может способствовать азотному циклу и увеличивать поглощение и использование азота растениями (Zeng et al., 2014). Следовательно, более высокая численность этих бактерий может улучшить устойчивость сои в ризосфере.Однако реакция этих родов на долгосрочное непрерывное земледелие требует дальнейшего изучения их функционального значения в ответ на различные системы долгосрочного непрерывного земледелия.
Был проведен анализ ассоциативной сети, чтобы получить более комплексное представление о составе бактериального сообщества и сравнить сложность сетей, действующих в ризосферных почвах, генотипов сои, устойчивых к непрерывному земледелию, и генотипов сои, чувствительных к непрерывному земледелию (Jiang et al., 2017). Сети двух генотипов по-разному реагировали на непрерывное выращивание. Модульность и отрицательные корреляции в сети толерантного генотипа положительно отреагировали на непрерывное земледелие, предполагая, что между доминантными бактериальными родами было больше событий сцепления, сотрудничества и обмена (Coyte et al., 2015). Для чувствительного генотипа отрицательная корреляция и средняя степень (avgK) увеличились в поле 7-летней системы непрерывного земледелия, но снизились в поле 36-летней системы непрерывного земледелия, что свидетельствует о нестабильности чувствительного генотипа и коротких периодах непрерывного посева. может иметь пагубное влияние на стабильность микробного сообщества, но этот эффект можно уменьшить, увеличив периоды непрерывного посева.Этот результат согласуется с предыдущим исследованием, в котором было обнаружено, что система долгосрочного непрерывного земледелия имеет микробные взаимодействия, аналогичные таковым в системе здорового севооборота (Liu et al., 2020). Кроме того, важные микробные таксоны в сетях, которые служат «движущими силами» от севооборотов к краткосрочному или продолжительному возделыванию, различались между двумя генотипами сои, что указывает на то, что ответ ризосферного микробного сообщества под разными генотипами сои на кратковременный и продолжительный -временное непрерывное земледелие было другим.
Таким образом, краткосрочное или долгосрочное непрерывное земледелие не оказало значительного влияния на бактериальное альфа-разнообразие ризосферы почвы. Кратковременный непрерывный посев увеличивал количество нематод цист сои, в то время как длительный непрерывный посев уменьшал это количество, а у толерантных генотипов было меньше нематод цист сои, чем у чувствительных генотипов. Кроме того, непрерывное земледелие значительно увеличило потенциально полезные популяции бактерий по сравнению с севооборотом и краткосрочным непрерывным возделыванием, что свидетельствует о том, что долгосрочное непрерывное возделывание сои сдвигает микробное сообщество в здоровую систему севооборота.Толерантные генотипы сои могут привлекать некоторые микроорганизмы, которые играют важную роль в устойчивости соевых бобов к длительному непрерывному возделыванию. Однако для подтверждения этих результатов необходимы дальнейшие эксперименты с изоляцией и проверкой видов. Более того, сети двух генотипов по-разному реагировали на непрерывное выращивание. Толерантный генотип положительно отреагировал на непрерывный посев, в то время как для чувствительного генотипа топология предполагала, что нестабильность чувствительного генотипа и короткие периоды непрерывной посадки могут иметь пагубный эффект, но этот эффект можно смягчить с увеличением периодов непрерывной посадки.
Заявление о доступности данныхНаборы данных, представленные в этом исследовании, можно найти в онлайн-репозиториях. Названия репозитория / репозиториев и номера доступа можно найти в статье / Дополнительных материалах.
Взносы авторов
TL, HN и MY разработали исследование. MY, KY, LW, DH и SW провели исследование. TY, QS, HX и RW проанализировали данные. MY и TL проанализировали данные и написали рукопись. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Эта работа была поддержана Китайской системой сельскохозяйственных исследований Министерства финансов и MARA (CARS-04), а также проектами независимых исследований лаборатории (SKLCUSA-a201913).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Примечание издателя
Все утверждения, выраженные в этой статье, принадлежат исключительно авторам и не обязательно отражают претензии их дочерних организаций или издателей, редакторов и рецензентов.Любой продукт, который может быть оценен в этой статье, или заявление, которое может быть сделано его производителем, не подлежат гарантии или одобрению со стороны издателя.
Благодарности
Мы хотели бы поблагодарить Чай Сяокуи и Цяо Чжисянь из Центра анализа и тестирования Института гидробиологии Китайской академии наук за их помощь в ПЦР-амплификации, секвенировании и анализе данных.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https: // www.frontiersin.org/articles/10.3389/fmicb.2021.729047/full#supplementary-material
Сноски
Список литературы
Agler, M. T., Ruhe, J., Kroll, S., Morhenn, C., Kim, S. T., Weigel, D., et al. (2016). Таксоны-концентраторы микробов связывают факторы хозяина и абиотические факторы с изменчивостью микробиома растений. PLoS Biol. 14: e1002352. DOI: 10.1371 / journal.pbio.1002352
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Авидано, Л., Гамалеро, Э., Косса, Г.П., и Карраро, Э. (2005). Характеристика здоровья почвы на итальянском загрязненном участке с использованием микроорганизмов в качестве биоиндикаторов. Заявл. Soil Ecol. 30, 21–33. DOI: 10.1016 / j.apsoil.2005.01.003
CrossRef Полный текст | Google Scholar
Бай, Л., Цуй, Дж. К., Цзе, В. Г., и Цай, Б. Ю. (2015). Анализ состава сообществ ризосферных грибов на полях сплошных посевов сои. Microbiol. Res. 180, 49–56. DOI: 10.1016 / j.micres.2015.07.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берри, Д., и Виддер, С. (2014). Расшифровка микробных взаимодействий и обнаружение ключевых видов с помощью сетей совместного возникновения. Фронт. Microbiol. 5: 219. DOI: 10.3389 / fmicb.2014.00219
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цай, Б. Ю., Ван, Л. Ю., Ху, В., Цзе, В. Г., и Лин, Х. З. (2015). Анализ структуры сообщества патогенных грибов корневых гнилей на всходовой стадии сплошного возделывания сои. Подбородок. Agr. Sci. Бык. 31, 92–98.
Google Scholar
Чиген Т., Мухаммад, А. Н., Аяка, К., Бао, Л., Чон, Д. Л., Ынхо, С. и др. (2018). Разнообразие профилей изофлавонов в корейских диких соевых бобах (Glycine soja Sieb. & Zucc.). Turk. J. Agric. Для. 42, 248–261. DOI: 10.3906 / tar-1709-119
CrossRef Полный текст | Google Scholar
Чонг, Дж., Лю, П., Чжоу, Г., и Ся, Дж. (2020). Использование MicrobiomeAnalyst для комплексного статистического, функционального и мета-анализа данных микробиома. Нат. Protoc. 5, 799–821. DOI: 10.1038 / s41596-019-0264-1
CrossRef Полный текст | Google Scholar
Коул, Дж. Р., Ван, К., Карденас, Э., Фиш, Дж., Чай, Б., Фаррис, Р. Дж. И др. (2009). Проект базы данных рибосом: улучшенное выравнивание и новые инструменты для анализа рРНК. Nucleic Acids Res. 37, 141–145. DOI: 10.1093 / nar / gkn879
CrossRef Полный текст | Google Scholar
Диас Т., Дьюкс А. и Антунес П. М. (2015). Учет биотического воздействия почвы на здоровье почвы и урожайность сельскохозяйственных культур при проектировании севооборотов. J. Sci. Продовольственное сельское хозяйство. 95, 447–454. DOI: 10.1002 / jsfa.6565
CrossRef Полный текст | Google Scholar
Эдгар Р. К., Хаас Б. Дж., Клементе Дж. К., Айва К. и Найт Р. (2011). UCHIME улучшает чувствительность и скорость обнаружения химер. Биоинформатика 27, 2194–2200. DOI: 10.1093 / биоинформатика / btr381
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Friendly, M., and Meyer, D. (2015). Анализ дискретных данных с помощью R: методы визуализации и моделирования категориальных данных и данных подсчета. Бока-Ратон: CRC Press, 63–108.
Google Scholar
Ху, Дж. К., и Ван, С. Дж. (1996). Исследование болезней почвы при непрерывном возделывании сои I. Влияние микотоксина, продуцируемого Penicillium purpurogenum . Подбородок. J. Appl. Ecol. 7, 396–400.
Google Scholar
Цзи, С. Н., Сяо, Ю. З., Тиан, Х. М., и Чжао, Ф. Х. (1996). Влияние стерилизованной почвы сои и стерни кукурузы на рост сои. Дж.К северо-востоку. Agr. 27, 326–329.
Google Scholar
Jiang, Y., Li, S., Li, R., Zhang, J., Liu, Y., Lv, L., et al. (2017). Сорта растений отражают состав ризосферного бактериального сообщества и сети ассоциаций. Soil Biol. Biochem. 109, 145–155. DOI: 10.1016 / j.soilbio.2017.02.010
CrossRef Полный текст | Google Scholar
Кайзерманн А., Фрис Ф. Т., Грифс Р. И. и Барджет Р. Д. (2017). Унаследованное влияние засухи на обратную связь между растениями и почвой и взаимодействие растений с растениями. New Phytol. 215, 1413–1424. DOI: 10.1111 / nph.14661
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кунтал, Б.К., Чандракар, П., Садху, С., и Манде, С.С. (2019). «NetShift»: методология понимания «возбудителей микробов» на основе наборов данных о здоровом и болезненном микробиоме. ISME J. 13, 442–454. DOI: 10.1038 / s41396-018-0291-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Квак, М. Дж., Конг, Х. Г., Чой, К., Kwon, S., Song, J. Y., Lee, J., et al. (2018). Структура микробиома ризосферы изменяется, чтобы обеспечить устойчивость томатов к увяданию. Нат. Biotechnol. 36, 1100–1109. DOI: 10.1038 / NBT.4232
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лебейс, С. Л., Паредес, С. Х., Лундберг, Д. С., Брейкфилд, Н., Геринг, Дж., Макдональд, М. и др. (2015). Салициловая кислота модулирует колонизацию корневого микробиома определенными таксонами бактерий. Наука 349, 860–864.DOI: 10.1126 / science.aaa8764
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли В., Годзик А. (2015). Cd-hit: быстрая программа для кластеризации и сравнения больших наборов белковых или нуклеотидных последовательностей. Биоинформатика 22, 1658–1659. DOI: 10.1007 / 978-1-4899-7478-5_221
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли, К. Г., Ли, Х. М., Конг, В. Д., Ву, Ю., и Ван, Дж. Г. (2010). Влияние монокультуры сои на микробное сообщество почвы в Северо-Восточном Китае. Почва растений 330, 423–433. DOI: 10.1007 / s11104-009-0216-6
CrossRef Полный текст | Google Scholar
Ли, Х. З., Руи, Дж. П., Мао, Й. Дж., Яннарелл, А., Маки, Р. (2014). Динамика структуры бактериального сообщества в ризосфере сорта кукурузы. Soil Biol. Biochem. 68, 392–401. DOI: 10.1016 / j.soilbio.2013.10.017
CrossRef Полный текст | Google Scholar
Лиан, Т. X., Ма, К. Б., Ши, К. Х., Цай, З. Д., Чжан, Ю. Ф., Cheng, Y.B., et al. (2019). Высокий алюминиевый стресс вызывает различную активность ферментов ризосферы в почве и структуру бактериального сообщества между устойчивыми к алюминию и чувствительными к нему генотипами сои. Почва растений 440, 409–425. DOI: 10.1007 / s11104-019-04089-8
CrossRef Полный текст | Google Scholar
Лю Х. Б., Буррас К. Л., Кравченко Ю. С., Дюран А., Хаффман Т., Моррас Х. и др. (2012). Обзор Mollisols в мире: распространение, землепользование и управление. Кан. J. Почвоведение. 92, 383–402. DOI: 10.4141 / cjss2010-058
CrossRef Полный текст | Google Scholar
Лю, З. X., Лю, Дж. Дж., Юа, З. Х., Яо, К., Ли, Ю. С., Лян, А. З. и др. (2020). Долгосрочные непрерывные посевы сои сравнимы с севооборотом по опосредованию численности, разнообразия и состава сообщества микробов. Обработка почвы Res. 197: 104503. DOI: 10.1016 / j.still.2019.104503
CrossRef Полный текст | Google Scholar
Лю, Х., Пан, Ф. Дж., Хан, Х. З., Сонг, Ф. Б., Чжан, З. М., Янь, Дж. И др. (2019). Реакция структуры почвенного грибного сообщества на длительный непрерывный посев сои. Фронт. Microbiol. 9: 3316. DOI: 10.3389 / fmicb.2018.03316
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мендес, Л. В., Мендес, Р., Рааймейкерс, Дж. М., и Цай, С. М. (2018). Селекция на устойчивость к почвенным патогенам влияет на активный ризосферный микробиом фасоли обыкновенной. ISME J. 12, 3038–3042.DOI: 10.1038 / s41396-018-0234-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мерилс, Дж. М., Гил, С. В., Конфорто, К., Фигони, Г., Ловера, Э., Марч, Г. Дж. И др. (2009). Сообщества почвенных микробов при различных системах возделывания сои: характеристика динамики микробной популяции, микробной активности почвы, микробной биомассы и профилей жирных кислот. Обработка почвы Res. 103, 271–281. DOI: 10.1016 / j.still.2008.10.008
CrossRef Полный текст | Google Scholar
Наутиял, К.С. (1999). Эффективная микробиологическая питательная среда для скрининга микроорганизмов, растворяющих фосфаты. FEMS Microbiol. Lett. 170, 265–270. DOI: 10.1111 / j.1574-6968.1999.tb13383.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перес-Брандан, К., Уидобро, Дж., Грюмберг, Б., Скандиани, М. М., Луке, А. Г., Мерилес, Дж. М. и др. (2014). Соевые грибковые болезни, передаваемые через почву: параметр для измерения влияния интенсификации сельского хозяйства на здоровье почвы. Кан. J. Microbiol. 60, 73–84. DOI: 10.1139 / cjm-2013-0792
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Филлипс Д., Фокс Т. К., Кинг М., Бхуванесвари Т. В. и Тойбер Л. Р. (2004). Микробные продукты вызывают выделение аминокислот из корней растений. Plant Physiol. 136, 2887–2894. DOI: 10.1104 / стр.104.044222
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ши, К. Х., Лю, Ю. Т., Ши, А. К., Цай, З.D., Nian, H., Hartmann, M., et al. (2020). Сообщества ризосферных почвенных грибов толерантных и чувствительных к алюминию генотипов сои по-разному реагируют на алюминиевый стресс в кислой почве. Фронт. Microbiol. 11: 1177. DOI: 10.3389 / fmicb.2020.01177
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сонг, Дж., Ли, С. X., Вэй, В., Сюй, Ю. Л. и Яо, К. (2017). Оценка паразитических грибов для уменьшения нематоды цист сои с подавляющей почвой на соевых полях Северо-Восточного Китая. Acta Agr. Сканд. B Почвенный завод 67, 730–736. DOI: 10.1080 / 0
10.2017.1343377CrossRef Полный текст | Google Scholar
Тан, Х., Сяо, К. Х., Ма, Дж. З., Ю, М., Ли, Ю. М., Ван, Г. Л. и др. (2009). Разнообразие прокариот в соевых почвах для непрерывного возделывания и севооборота. FEMS Microbiol. Lett. 298, 267–273. DOI: 10.1111 / j.1574-6968.2009.01730.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вентер, З.С., Джейкобс, К., Хокинс, Х. Дж. (2016). Влияние севооборота на микробное разнообразие почвы: метаанализ. Pedobiologia 59, 215–223. DOI: 10.1016 / j.pedobi.2016.04.001
CrossRef Полный текст | Google Scholar
Уолдроп, М. П., Бальзер, Т. К., и Файерстоун, М. К. (2000). Привязка состава микробного сообщества к функционированию в тропической почве. Soil Biol. Biochem. 32, 1837–1846. DOI: 10.1016 / S0038-0717 (00) 00157-7
CrossRef Полный текст | Google Scholar
Вэй, W., Сюй, Ю. Л., Ли, С. X., Чжу, Л., и Сун, Дж. (2015). Разработка почвы для подавления корневых болезней сои с непрерывным многолетним выращиванием сои в черноземах Северо-Восточного Китая. Acta Agr. Сканд. B Почвенный завод 65, 279–285. DOI: 10.1080 / 0
10.2014.992941CrossRef Полный текст | Google Scholar
Сюй, Ю. Л., Ван, Г. Х., и Хань, Х. З. (1995). Взаимосвязь между характеристиками экологического распределения почвенных микробов и заболеванием корней сои при монокультурном выращивании и севообороте сои. Syst. Sciemces Compr. Stud. Agr. 11, 311–314.
Google Scholar
Янь, М. К., Сюй, Т. Т., Сонг, П. Х. и Дай, Дж. Дж. (2012). Влияние различных моделей выращивания проростков сои и кукурузы на ферментативную активность почвы и МБК и МБН. J. Северо-восток. Agr. Univ. 19, 42–47. DOI: 10.1016 / S1006-8104 (13) 60049-5
CrossRef Полный текст | Google Scholar
Цзэн В., Бай X., Чжан Л., Ван А. и Пэн Ю. (2014). Динамика популяций нитрифицирующих бактерий для нитритации, достигнутая в процессе очистки городских сточных вод в Йоханнесбурге (JHB). Биоресурсы. Technol. 162, 30–37. DOI: 10.1016 / j.biortech.2014.03.102
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжан, X. М., Хань, X. Р., Ян, Дж. Ф. и Гао, З. К. (2004). Влияние посевов и стерней сои на экссудаты корней сои. Подбородок. J. Почвоведение. 35, 631–635. DOI: 10.1300 / J064v24n01_09
CrossRef Полный текст | Google Scholar
Чжоу, X. G., Wang, Z. L., Jia, H. T., Li, L., and Wu, F.З. (2018). Постоянно выращиваемый топинамбур изменил состав почвенного бактериального сообщества и численность бактерий, окисляющих и денитрифицирующих аммиак. Фронт. Microbiol. 9: 705. DOI: 10.3389 / fmicb.2018.00705
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжу, Ю. Б., Ши, Ф. Ю., Чжан, Р. Дж., И Ву, Ю. П. (2014). Сравнение бактериального разнообразия в почвах севооборотов и сплошных посевов сои в провинции Хэйлунцзян. Acta Phytophy. Грех. 41, 403–409.
Google Scholar
Zhu, L., Zeng, C.L., Li, Y.Q., Yu, B.Q., Gao, F., Wei, W., et al. (2017). Характеристика разнообразия бактериального сообщества на соевом поле при непрерывном возделывании на основе высокопроизводительного секвенирования. Soybean Sci. 36, 419–424.
Google Scholar
PFU Limited
- Японский
- ДОМ
- Информация о компании
- Направления деятельности
- Корпоративный профиль
- Сообщение
- Обещание бренда
- История
- Путь PFU
- ISO
- Финансовые результаты
- Политика закупок
- Окружающая среда
- Компании Группы ПФУ
- Штаб-квартира, центры исследований и разработок
- Филиалы сбыта
- Сервисные отделения
- Группа PFU (Внутренний)
- Группа PFU (за рубежом)
Меню
Продукты
Оборудование
- fi / SP Series / ScanSnap Series Сканеры изображений США / Европа / Китай
Восточная Азия / Япония - MEDIASTAFF Series Интерактивные киоски
- Встроенные продукты Встроенные компьютеры
- Happy Hacking Keyboard Компактные и усовершенствованные клавиатуры США / Европа / Китай / Япония
- REALFORCE R2 [PFU Limited Edition] Высококачественные клавиатуры без ключа США / Европа / Китай
Программное обеспечение
- Программное обеспечение DynaEye Series OCR для профессионального использования
- Серия BIP Программное обеспечение Business Form
Безопасность
- iNetSec Повышенная кибербезопасность
Решения и услуги
Интеграция услуг Кибербезопасность ИТ-услугиКорпоративная информация
Информация о компании
- Направления деятельности
- Корпоративный профиль
- Сообщение
- Обещание бренда
- История
- Путь PFU
- ISO
- Финансовые результаты
- Политика закупок
- Окружающая среда
- ДОМ
- Карта сайта
- Контакт
- Политика конфиденциальности